

- 1.22 MB
- 2022-04-22 11:26:36 发布


- 1、本文档共5页,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,可选择认领,认领后既往收益都归您。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细先通过免费阅读内容等途径辨别内容交易风险。如存在严重挂羊头卖狗肉之情形,可联系本站下载客服投诉处理。
- 文档侵权举报电话:19940600175。
'第二章遗传的细胞学基础 本章习题 1.解释下列名词:原核细胞、真核细胞、染色体、染色单体、着丝点、细胞周期、同源染色体、异源染色体、无丝分裂、有丝分裂、单倍体、二倍体、联会、胚乳直感、果实直感。答:原核细胞:一般较小,统称为原核生物。真核细胞:比原核细胞大,其结构和功能也比原核细胞复杂。真核细胞含有核物质和核结构,细胞核是遗传物质集聚的主要场所,对控制细胞发育和性状遗传起主导作用。另外真核细胞还含有线粒体、叶绿体、内质网等各种膜包被的细胞器。真核细胞都由细胞膜与外界隔离,细胞内有起支持作用的细胞骨架。染色体:含有许多基因的自主复制核酸分子。细菌的全部基因包容在一个双股环形DNA构成的染色体内。真核生物染色体是与组蛋白结合在一起的线状DNA双价体;整个基因组分散为一定数目的染色体,每个染色体都有特定的形态结构,染色体的数目是物种的一个特征。 染色单体:由染色体复制后并彼此靠在一起,由一个着丝点连接在一起的姐妹染色体。着丝点:在细胞分裂时染色体被纺锤丝所附着的位置。一般每个染色体只有一个着丝点,少数物种中染色体有多个着丝点,着丝点在染色体的位置决定了染色体的形态。细胞周期:包括细胞有丝分裂过程和两次分裂之间的间期。其中有丝分裂过程分为: (1)DNA合成前期(G1期);(2)DNA合成期(S期); (3)DNA合成后期(G2期);(4)有丝分裂期(M期)。 同源染色体:生物体中,形态和结构相同的一对染色体。 异源染色体:生物体中,形态和结构不相同的各对染色体互称为异源染色体。 无丝分裂:也称直接分裂,只是细胞核拉长,缢裂成两部分,接着细胞质也分裂,从而成为两个细胞,整个分裂过程看不到纺锤丝的出现。在细胞分裂的整个过程中,不象有丝分裂那样经过染色体有规律和准确的分裂。 有丝分裂:包含两个紧密相连的过程:核分裂和质分裂。即细胞分裂为二,各含有一个核。分裂过程包括四个时期:前期、中期、后期、末期。在分裂过程中经过染色体有规律的和准确的分裂,而且在分裂中有纺锤丝的出现,故称有丝分裂。 单倍体:具有一组基本染色体数的细胞或者个体。 二倍体:具有两组基本染色体数的细胞或者个体。 联会:减数分裂中,同源染色体的配对过程。 胚乳直感:植物经过了双受精,胚乳细胞是3n,其中2n来自极核,n来自精核,如果在3n胚乳的性状上由于精核的影响而直接表现父本的某些性状,这种现象称为胚乳直感。 果实直感:植物的种皮或果皮组织在发育过程中由于花粉影响而表现父本的某些性状,称为果实直感。
2.细胞的膜体系包括哪些膜结构?细胞质里包括哪些主要的细胞器?各有什么特点? 答:细胞的膜体系包括膜结构有:细胞膜、线粒体、质体、内质网、高尔基体、液泡、核膜。 细胞质里主要细胞器有:线粒体、叶绿体、核糖体、内质网、中心体。各细胞器特点如下: 线粒体:两层膜组成,线粒体含有多种氧化酶,能进行氧化磷酸化,可传递和贮存所产生的能量,成为细胞里氧化作用和呼吸作用的中心,是细胞的动力工厂。线粒体含有DNA、RNA和核糖体,具有独立合成蛋白质的能力。有独立的遗传体系。 叶绿体:双层膜,叶绿体含有DNA、RNA及核糖体等,能够合成蛋白质并且能够分裂增殖。这些特征都表明叶绿体具有特定的遗传功能,是遗传物质的载体之一。 核糖体:无膜,数量多。核糖体是由蛋白质和RNA组成。已知核糖体是合成蛋白质的主要场所。 内质网:单层膜。把质膜和核膜连成一个完整膜体系,为细胞空间提供了支架作用,是蛋白质合成的主要场所,并通过内质网将合成的蛋白质运送到细胞的其它部位。平滑内质网,可能与某些激素合成有关。 中心体:中心体是动物和某些蕨类及裸子植物细胞特有的细胞器。其含有一对由微管蛋白组成的结构复杂的中心粒。它与细胞的有丝分裂和减数分裂过程中纺锤丝的形成有关。 3.一般染色体的外部形态包括哪些部分?染色体形态有哪些类型? 答:一般染色体的外部形态包括:着丝粒、染色体两个臂、主溢痕、次溢痕、随体。 一般染色体的类型有:V型、L型、棒型、颗粒型。 4.植物的10个花粉母细胞可以形成:多少花粉粒?多少精核?多少管核?又10个卵母细胞可以形成:多少胚囊?多少卵细胞?多少极核?多少助细胞?多少反足细胞? 答:植物的10个花粉母细胞可以形成: 花粉粒:10×4=40个;精核:40×2=80个;管核:40×1=40个。 10个卵母细胞可以形成: 胚囊:10×1=10个;卵细胞:10×1=10个;极核:10×2=20个; 助细胞:10×2=20个;反足细胞:10×3=30个。
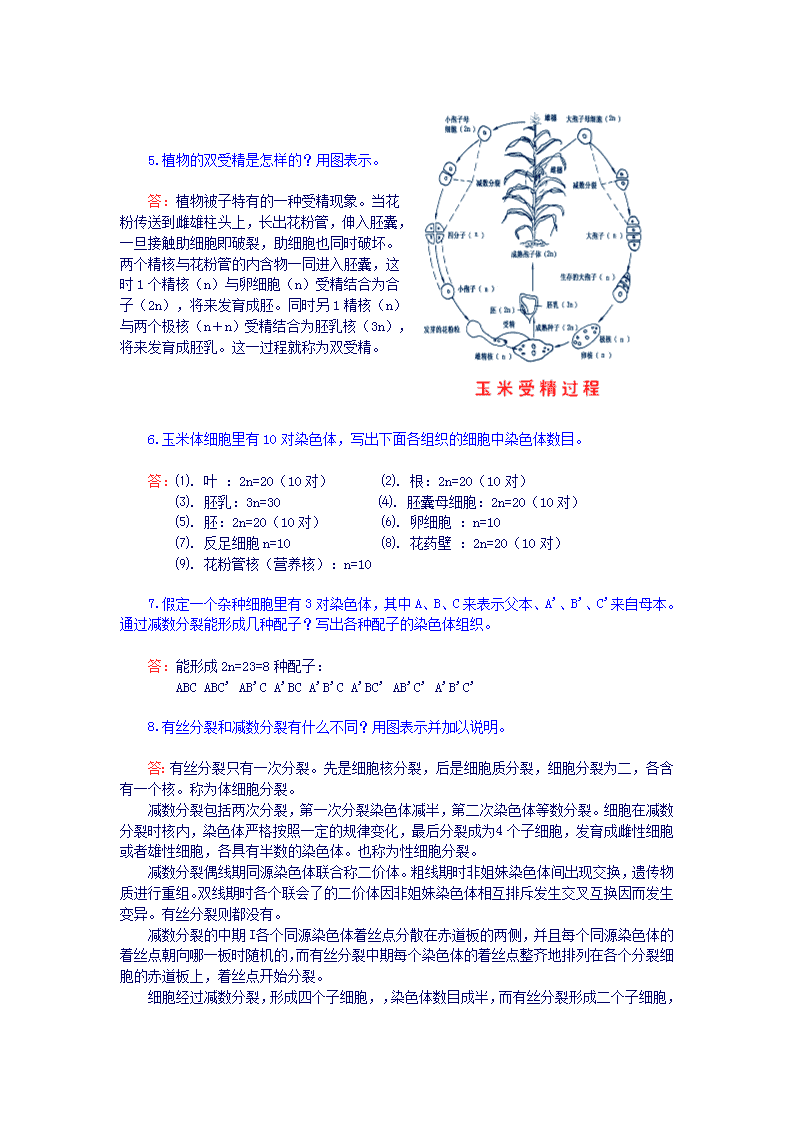
5.植物的双受精是怎样的?用图表示。 答:植物被子特有的一种受精现象。当花粉传送到雌雄柱头上,长出花粉管,伸入胚囊,一旦接触助细胞即破裂,助细胞也同时破坏。两个精核与花粉管的内含物一同进入胚囊,这时1个精核(n)与卵细胞(n)受精结合为合子(2n),将来发育成胚。同时另1精核(n)与两个极核(n+n)受精结合为胚乳核(3n),将来发育成胚乳。这一过程就称为双受精。 6.玉米体细胞里有10对染色体,写出下面各组织的细胞中染色体数目。 答:⑴.叶:2n=20(10对) ⑵.根:2n=20(10对) ⑶.胚乳:3n=30 ⑷.胚囊母细胞:2n=20(10对) ⑸.胚:2n=20(10对) ⑹.卵细胞:n=10 ⑺.反足细胞n=10 ⑻.花药壁:2n=20(10对) ⑼.花粉管核(营养核):n=10 7.假定一个杂种细胞里有3对染色体,其中A、B、C来表示父本、A"、B"、C"来自母本。通过减数分裂能形成几种配子?写出各种配子的染色体组织。 答:能形成2n=23=8种配子: ABCABC"AB"CA"BCA"B"CA"BC"AB"C"A"B"C" 8.有丝分裂和减数分裂有什么不同?用图表示并加以说明。 答:有丝分裂只有一次分裂。先是细胞核分裂,后是细胞质分裂,细胞分裂为二,各含有一个核。称为体细胞分裂。 减数分裂包括两次分裂,第一次分裂染色体减半,第二次染色体等数分裂。细胞在减数分裂时核内,染色体严格按照一定的规律变化,最后分裂成为4个子细胞,发育成雌性细胞或者雄性细胞,各具有半数的染色体。也称为性细胞分裂。 减数分裂偶线期同源染色体联合称二价体。粗线期时非姐妹染色体间出现交换,遗传物质进行重组。双线期时各个联会了的二价体因非姐妹染色体相互排斥发生交叉互换因而发生变异。有丝分裂则都没有。 减数分裂的中期I
各个同源染色体着丝点分散在赤道板的两侧,并且每个同源染色体的着丝点朝向哪一板时随机的,而有丝分裂中期每个染色体的着丝点整齐地排列在各个分裂细胞的赤道板上,着丝点开始分裂。 细胞经过减数分裂,形成四个子细胞,,染色体数目成半,而有丝分裂形成二个子细胞,染色体数目相等。 9.有丝分裂和减数分裂意义在遗传学上各有什么意义在遗传学上? 答:有丝分裂在遗传学上的意义:多细胞生物的生长主要是通过细胞数目的增加和细胞体积的增大而实现的,所以通常把有丝分裂称为体细胞分裂,这一分裂方式在遗传学上具有重要意义。首先是核内每个染色体准确地复制分裂为二,为形成两个在遗传组成上与母细胞完全一样的子细胞提供了基础。其次是复制后的各对染色体有规则而均匀地分配到两个子细胞中去,使两个细胞与母细胞具有同样质量和数量的染色体。对细胞质来说,在有丝分裂过程中虽然线粒体、叶绿体等细胞器也能复制、增殖数量。但是它们原先在细胞质中分布是不恒定的,因而在细胞分裂时它们是随机而不均等地分配到两个细胞中去。由此可见,任何由线粒体、叶绿体等细胞器所决定的遗传表现,是不可能与染色体所决定的遗传表现具有同样的规律性。这种均等方式的有丝分裂既维持了个体的正常生长和发育,也保证了物种的连续性和稳定性。植物采用无性繁殖所获得的后代能保持其母本的遗传性状,就在于它们是通过有丝分裂而产生的。减数分裂在遗传学上的意义:在生物的生活周期中,减数分裂是配子形成过程中的必要阶段。这一分裂方式包括两次分裂,其中第二次分裂与一般有丝分裂基本相似;主要是第一次分裂是减数的,与有丝分裂相比具有明显的区别,这在遗传学上具有重要的意义。首先,减数分裂时核内染色体严格按照一定规律变化,最后经过两次连续的分裂形成四个子细胞,发育为雌雄性细胞,但遗传物质只进行了一次复制,因此,各雌雄性细胞只具有半数的染色体(n)。这样雌雄性细胞受精结合为合子,又恢复为全数的染色体(2n),从而保证了亲代与子代之间染色体数目的恒定性,为后代的正常发育和性状遗传提供了物质基础;同时保证了物种相对的稳定性。其次,各对同源染色体在减数分裂中期I排列在赤道板上,然后分别向两极拉开,各对染色体中的两个成员在后期I分向两极时是随机的,即一对染色体的分离与任何另一对染色体的分离不发生关联,各个非同源染色体之间均可能自由组合在一个子细胞里。n对染色体,就可能有2n种自由组合方式。例如,水稻n=12,其非同源染色体分离时的可能组合数既为212
=4096。这说明各个细胞之间在染色体上将可能出现多种多样的组合。不仅如此,同源染色体的非姐妹染色单体之间的片段还可能出现各种方式的交换,这就更增加了这种差异的复杂性。因而为生物的变异提供的重要的物质基础,有利于生物的适应及进化,并为人工选择提供了丰富的材料。 10.何谓无融合生殖?它包含有哪几种类型? 答:无融合生殖是指雌雄配子不发生核融合的一种无性生殖方式,被认为是有性生殖的一种特殊方式或变态。 它有以下几种类型: ⑴.营养的无融合生殖; ⑵.无融合结子:包括①.单倍配子体无融合生殖;②.二倍配子体无融合生殖;③.不定胚; ⑶.单性结实。 11.以红色面包霉为例说明低等植物真菌的生活周期,它与高等植物的生活周期有何异同? 答:红色面包霉的单倍体世代(n=7)是多细胞的菌丝体和分生孢子。由分生孢子发芽形成为新的菌丝,属于其无性世代。一般情况下,它就是这样循环地进行无性繁殖。但是,有时也会产生两种不同生理类型的菌丝,一般分别假定为正(+)和(-)两种结合型,它们将类似于雌雄性别,通过融合和异型核的接合而形成二倍体的合子(2n=14),属于其有性世代。合子本身是短暂的二倍体世代。红色面包霉的有性过程也可以通过另一种方式来实现。因为其"+"和"-"两种接合型的菌丝都可以产生原子囊果和分生孢子。如果说原子囊果相当于高等植物的卵细胞,则分生孢子相当于精细胞。这样当"+"接合型(n)与"-"接合型(n)融合和受精后,便可形成二倍体的合子(2n)。无论上述的那一种方式,在子囊果里子囊的菌丝细胞中合子形成以后,可立即进行两次减数分裂(一次DNA复制和二次核分裂),产生出四个单倍体的核,这时称为四个孢子。四个孢子中每个核进行一次有丝分裂,最后形成为8个子囊孢子,这样子囊里的8个孢子有4个为"+"接合型,另有4个为"-"接合型,二者总是成1:1的比例分离。 低等植物和高等植物的一个完整的生活周期,都是交替进行着无性世代和有性世代。它们都具有自己的单倍体世代和二倍体世代,只是低等植物的世代的周期较短(它的有性世代可短到10天),并且能在简单的化学培养基上生长。而高等植物的生活周期较长,配子体世代孢子体世代较长,繁殖的方式和过程都是高等植物比低等植物复杂得多。 12.高等植物与高等动物的生活周期有什么主要差异?用图说明。 答:
高等动、植物生活周期的主要差异:动物通常是从二倍体的性原细胞经过减数分裂即直接形成精子和卵细胞,其单倍体的配子时间很短;有性过程是精子和卵细胞融合成受精卵,再由受精卵分化发育成胚胎,直至成熟个体。而植物从二倍体的性原细胞经过减数分裂后先产生为单倍体的雄配子体和雌配子体,再进行一系列的有丝分裂,然后再形成为精子和卵细胞;有性过程是经双受精,精子与卵细胞结合进一步发育分化成胚,而另一精子与两个极核结合,发育成胚乳,胚乳在胚或种子生长发育过程起到很重要作用。具体差异见下图:
第三章遗传物质的分子基础 本章习题 1.解释下列名词:半保留复制、冈崎片段、转录、翻译、小核RNA、不均一核RNA、遗传密码简并、多聚核糖体、中心法则。 半保留复制:DNA分子的复制,首先是从它的一端氢键逐渐断开,当双螺旋的一端已拆开为两条单链时,各自可以作为模板,进行氢键的结合,在复制酶系统下,逐步连接起来,各自形成一条新的互补链,与原来的模板单链互相盘旋在一起,两条分开的单链恢复成DNA双分子链结构。这样,随着DNA分子双螺旋的完全拆开,就逐渐形成了两个新的DNA分子,与原来的完全一样。这种复制方式成为半保留复制。 冈崎片段:在DNA复制叉中,后随链上合成的DNA不连续小片段称为冈崎片段。 转录:由DNA为模板合成RNA的过程。RNA的转录有三步: ①.RNA链的起始; ②.RNA链的延长; ③.RNA链的终止及新链的释放。 翻译:以RNA为模版合成蛋白质的过程即称为遗传信息的翻译过程。 小核RNA:是真核生物转录后加工过程中RNA的剪接体的主要成分,属于一种小分子RNA,可与蛋白质结合构成核酸剪接体。 不均一核RNA:在真核生物中,转录形成的RNA中,含有大量非编码序列,大约只有25%RNA经加工成为mRNA,最后翻译为蛋白质。因为这种未经加工的前体mRNA在分子大小上差别很大,所以称为不均一核RNA。 遗传密码:是核酸中核苷酸序列指定蛋白质中氨基酸序列的一种方式,是由三个核苷酸组成的三联体密码。密码子不能重复利用,无逗号间隔,存在简并现象,具有有序性和通用性,还包含起始密码子和终止密码子。 简并:一个氨基酸由一个以上的三联体密码所决定的现象。 多聚合糖体:一条mRNA分子可以同时结合多个核糖体,形成一串核糖体,成为多聚核糖体。 中心法则:蛋白质合成过程,也就是遗传信息从DNA-mRNA-蛋白质的转录和翻译的过程,以及遗传信息从DNA到DNA的复制过程,这就是生物学的中心法则。 2.如何证明DNA是生物的主要遗传物质? 答:
DNA作为生物的主要遗传物质的间接证据: ⑴.每个物种不论其大小功能如何,其DNA含量是恒定的。 ⑵.DNA在代谢上比较稳定。⑶.基因突变是与DNA分子的变异密切相关的。 DNA作为生物的主要遗传物质的直接证据: ⑴.细菌的转化已使几十种细菌和放线菌成功的获得了遗传性状的定向转化,证明起转化作用的是DNA; ⑵.噬菌体的侵染与繁殖主要是由于DNA进入细胞才产生完整的噬菌体,所以DNA是具有连续性的遗传物质。 ⑶.烟草花叶病毒的感染和繁殖说明在不含DNA的TMV中RNA就是遗传物质。 3.简述DNA双螺旋结构及其特点? 答:根据碱基互补配对的规律,以及对DNA分子的X射线衍射研究的成果,提出了DNA双螺旋结构。 特点:⑴.两条多核苷酸链以右手螺旋的形式,彼此以一定的空间距离,平行的环绕于同一轴上,很像一个扭曲起来的梯子。⑵.两条核苷酸链走向为反向平行。⑶.每条长链的内侧是扁平的盘状碱基。⑷.每个螺旋为3.4nm长,刚好有10个碱基对,其直径为2nm。⑸.在双螺旋分子的表面有大沟和小沟交替出现。 4.比较A-DNA、B-DNA、Z-DNA的主要异同? 答:A-DNA是DNA的脱水构型,也是右手螺旋,但每螺旋含有11个核苷酸对。比较短和密,其平均直径是2.3nm。大沟深而窄,小沟宽而浅。在活体内DNA并不以A构型存在,但细胞内DNA-RNA或RNA-RNA双螺旋结构,却与A-DNA非常相似。 B-DNA是DNA在生理状态下的构型。生活细胞中极大多数DNA以B-DNA形式存在。但当外界环境条件发生变化时,DNA的构型也会发生变化。 Z-DNA是某些DNA序列可以以左手螺旋的形式存在。当某些DNA序列富含G-C,并且在嘌呤和嘧啶交替出现时,可形成Z-DNA。其每螺旋含有12个核苷酸对,平均直径是1.8nm,并只有一个深沟。现在还不清楚Z-DNA在体内是否存在。 5.染色质的基本结构是什么?现有的假说是怎样解释染色质螺旋化为染色体的? 答:染色质是染色体在细胞分裂的间期所表现的形态,呈纤细的丝状结构,故也称染色质线。其基本结构单位是核小体、连接体和一个分子的组蛋白H1。每个核小体的核心是由H2A、H2B、H3和H4四种组蛋白各以两个分子组成的八聚体,其形状近似于扁球状。DNA双螺旋就盘绕在这八个组蛋白分子的表面。连接丝把两个核小体串联起来,是两个核小体之间的DNA双链。 细胞分裂过程中染色线卷缩成染色体:现在认为至少存在三个层次的卷缩:第一个层次是DNA分子超螺旋转化形成核小体,产生直径为10nm的间期染色线,在此过程中组蛋白H2A、H2B、H3和H4参与作用。第二个层次是核小体的长链进一步螺旋化形成直径为30nm的超微螺旋,称为螺线管,在此过程中组蛋白H1起作用。最后是染色体螺旋管进一步卷缩,并附着于由非组蛋白形成的骨架或者称中心上面成为一定形态的染色体。 6.原核生物DNA聚合酶有哪几种?各有何特点? 答:
原核生物DNA聚合酶有DNA聚合酶I、DNA聚合酶II和DNA聚合酶III。 DNA聚合酶I:具有5"-3"聚合酶功能外,还具有3"-5"核酸外切酶和5"-3"核酸外切酶的功能。 DNA聚合酶II:是一种起修复作用的DNA聚合酶,除具有5"-3"聚合酶功能外,还具有3"-5"核酸外切酶,但无5"-3"外切酶的功能。 DNA聚合酶III:除具有5"-3"聚合酶功能外,也有3"-5"核酸外切酶,但无3"-5"外切酶的功能。 7.真核生物与原核生物DNA合成过程有何不同? 答:⑴.真核生物DNA合成只是发生在细胞周期中的S期,原核生物DNA合成过程在整个细胞生长期中均可进行。 ⑵.真核生物染色体复制则为多起点的,而原核生物DNA复制是单起点的。 ⑶.真核生物DNA合成所需的RNA引物及后随链上合成的冈崎片段的长度比原核生物的要短。⑷.在真核生物中,有α、β、γ、δ和ε5种DNA聚合酶,δ是DNA合成的主要酶,由DNA聚合酶α控制后随链的合成,而由DNA聚合酶δ控制前导链的合成。既在真核生物中,有两种不同的DNA聚合酶分别控制前导链和后随链的合成。在原核生物DNA合成过程中,有DNA聚合酶I,DNA聚合酶II和DNA聚合酶III,并由DNA聚合酶III同时控制两条链的合成。⑸.真核生物的染色体为线状,有染色体端体的复制,而原核生物的染色体大多数为环状。 8.简述原核生物RNA的转录过程。 答:RNA的转录有三步: ⑴.RNA链的起始:首先是RNA聚合酶在δ因子的作用下结合于DNA的启动子部位,并在RNA聚合酶的作用下,使DNA双链解开,形成转录泡,为RNA合成提供单链模板,并按照碱基配对的原则,结合核苷酸,然后,在核苷酸之间形成磷酸二脂键,使其相连,形成RNA新链。δ因子在RNA链伸长到8-9个核苷酸后被释放,然后由核心酶催化RNA链的延长。 ⑵.RNA链的延长:RNA链的延长是在δ因子释放以后,在RNA聚合酶四聚体核心酶催化下进行。因RNA聚合酶同时具有解开DNA双链,并使其重新闭合的功能。随着RNA链的延长,RNA聚合酶使DNA双链不断解开和闭合。RNA转录泡也不断前移,合成新的RNA链。⑶.RNA链的终止及新链的释放:当RNA链延伸到终止信号时,RNA转录复合体就发生解体,而使新合成的RNA链得以释放。 9.真核生物与原核生物相比,其转录过程有何特点? 答:真核生物转录的特点: ⑴.在细胞核内进行。 ⑵.mRNA分子一般只编码一个基因。 ⑶.RNA聚合酶较多。 ⑷.RNA聚合酶不能独立转录RNA。 原核生物转录的特点: ⑴.原核生物中只有一种RNA聚合酶完成所有RNA转录。 ⑵.一个mRNA分子中通常含有多个基因。
10.简述原核生物蛋白质合成的过程。 答:蛋白质的合成分为链的起始、延伸和终止阶段: 链的起始:不同种类的蛋白质合成主要决定于mRNA的差异。在原核生物中,蛋白质合成的起始密码子为AUG。编码甲酰化甲硫氨酸。蛋白质合成开始时,首先是决定蛋白质起始的甲酰化甲硫氨酰tRNA与起始因子IF2结合形成第一个复合体。同时,核糖体小亚基与起始因子IF3和mRNA结合形成第二个复合体。接着两个复合体在始因子IF1和一分子GDP的作用下,形成一个完整的30S起始复合体。此时,甲酰化甲硫氨酰tRNA通过tRNA的反密码子识别起始密码AUG,而直接进入核糖体的P位(peptidyl,P)并释放出IF3。最后与50S大亚基结合,形成完整的70核糖体,此过程需要水解一分子GDP以提供能量,同时释放出IF1和IF2,完成肽链的起始。 链的延伸:根据反密码子与密码子配对的原则,第二个氨基酰tRNA进入A位。随后在转肽酶的催化下,在A位的氨基酰tRNA上的氨基酸残基与在P位上的氨基酸的碳末端间形成多肽键。此过程水解与EF-Tu结合的GTP而提供能量。最后是核糖体向前移一个三联体密码,原来在A位的多肽tRNA转入P位,而原在P的tRNA离开核糖体。此过程需要延伸因子G(EF-G)和水解GTP提供能量。这样空出的A位就可以接合另一个氨基酰tRNA,从而开始第二轮的肽链延伸。 链的终止:当多肽链的延伸遇到UAAUAGUGA等终止密码子进入核糖体的A位时,多肽链的延伸就不再进行。对终止密码子的识别,需要多肽释放因子的参与。在大肠杆菌中有两类释放因子RF1和RF2,RF1识别UAA和UAG,RF2识别UAA和UGA。在真核生物中只有释放因子eRF,可以识别所有三种终止密码子。
第四章孟德尔遗传 本章习题 1.小麦毛颖基因P为显性,光颖基因p为隐性。写出下列杂交组合的亲本基因型: (1)毛颖×毛颖,后代全部毛颖。 (2)毛颖×毛颖,后代3/4为毛颖1/4光颖。 (3)毛颖×光颖,后代1/2毛颖1/2光颖。 答:(1)亲本基因型为:PP×PP;PP×Pp; (2)亲本基因型为:Pp×Pp; (3)亲本基因型为:Pp×pp。 2.小麦无芒基因A为显性,有芒基因a为隐性。写出下列个各杂交组合中F1的基因型和表现型。每一组合的F1群体中,出现无芒或有芒个体的机会是多少? (1)AA×aa, (2)AA×Aa, (3)Aa×Aa, (4)Aa×aa, (5)aa×aa, 答:⑴.F1的基因型:Aa;F1的表现型:全部为无芒个体。 ⑵.F1的基因型:AA和Aa;F1的表现型:全部为无芒个体。 ⑶.F1的基因型:AA、Aa和aa;F1的表现型:无芒:有芒=3:1。 ⑷.F1的基因型:Aa和aa;F1的表现型:无芒河忻?1:1。 ⑸.F1的基因型:aa;F1的表现型:全部有芒个体。 3.小麦有稃基因H为显性,裸粒基因h为隐性。现以纯合的有稃品种(HH)与纯合的裸粒品种(hh)杂交,写出其F1和F2的基因型和表现型。在完全显性的条件下,其F2基因型和表现型的比例怎么样? 答:F1的基因型:Hh,F1的表现型:全部有稃。 F2的基因型:HH:Hh:hh=1:2:1,F2的表现型:有稃:无稃=3:1 4.大豆的紫花基因P对白花基因p为显性,紫花×白花的F1全为紫花,F2共有1653株,其中紫花1240株,白花413株,试用基因型说明这一试验结果。 答:由于紫花×白花的F1全部为紫花:即基因型为:PP×pp?Pp。 而F2基因型为:Pp×Pp?PP:Pp:pp=1:2:1,共有1653株,且紫花:白花=1240:413=3:1,符合孟得尔遗传规律。 5.纯种甜玉米和纯种非甜玉米间行种植,收获时发现甜粒玉米果穗上结有非甜玉米的子实,而非甜玉米果穗上找不到甜粒的子实,如何解释这一现象?怎么样验证解释? 答:
⑴.为胚乳直感现象,在甜粒玉米果穗上有的子粒胚乳由于精核的影响而直接表现出父本非甜显性特性的子实。原因:由于玉米为异花授粉植物,间行种植出现互相授粉,并说明甜粒和非甜粒是一对相对性状,且非甜粒为显性性状,甜粒为隐性性状(假设A为非甜粒基因,a为甜粒基因)。 ⑵.用以下方法验证: 测交法:将甜粒玉米果穗上所结非甜玉米的子实播种,与纯种非甜玉米测交,其后代的非甜粒和甜粒各占一半,既基因型为:Aa×aa=1:1,说明上述解释正确。 自交法:将甜粒玉米果穗上所结非甜玉米的子实播种,使该套袋自交,自交后代性状比若为3:1,则上述解释正确。 6.花生种皮紫色(R)对红色(r)为显性,厚壳T对薄壳t为显性。R-r和T-t是独立遗传的。指出下列各种杂交组合的:1.亲本基因型、配子种类和比例。2.F1的基因型种类和比例、表现型种类和比例。 答:祥见下表:杂交基因型亲本表现型配子种类配子比例F1基因型F1表现型TTrr×ttRR厚壳红色薄壳紫色Tr:tR1:1TtRr厚壳紫色TTRR×ttrr厚壳紫色薄壳红色TR:tr1:1TtRr厚壳紫色TtRr×ttRr厚壳紫色薄壳紫色TR:tr:tR:Tr1:3:3:1TtRR:ttRr:TtRr:ttRR:Ttrr:ttrr=1:2:2:1:1:1厚壳紫色:薄壳紫色:厚壳红色:薄壳红色=3:3:1:1ttRr×Ttrr薄壳紫色厚壳红色tR:tr:Tr1:2:1TtRr:Ttrr:ttRr:ttrr=1:1:1:1厚壳紫色:厚壳红色:薄壳紫色:薄壳红色=1:1:1:1 7.番茄的红果Y对黄果y为显性,二室M对多室m为显性。两对基因是独立遗传的。当一株红果二室的番茄与一株红果多室的番茄杂交后,F1群体内有3/8的植株为红果二室的,3/8是红果多室的,1/8是黄果二室的,1/8是黄果多室的。试问这两个亲本植株是怎样的基因型? 答:番茄果室遗传:二室M对多室m为显性,其后代比例为: 二室:多室=(3/8+1/8):(3/8+1/8)=1:1,因此其亲本基因型为:Mm×mm。 番茄果色遗传:红果Y对黄果y为显性,其后代比例为: 红果:黄果=(3/8+3/8):(1/8+1/8)=3:1, 因此其亲本基因型为:Yy×Yy。因为两对基因是独立遗传的,所以这两个亲本植株基因型:YyMm×Yymm。 8.下表是不同小麦品种杂交后代产生的各种不同表现性的比例,试写出各个亲本基因型(设毛颖、抗锈为显性)。
亲本组合毛颖抗锈毛颖感锈光颖抗锈光颖感锈毛颖感锈×光颖感锈毛颖抗锈×光颖感锈毛颖抗锈×光颖抗锈光颖抗锈×光颖抗锈01015018870081632149512 答:根据其后代的分离比例,得到各个亲本的基因型: (1)毛颖感锈×光颖感锈:Pprr×pprr (2)毛颖抗锈×光颖感锈:PpRr×pprr (3)毛颖抗锈×光颖抗锈:PpRr×ppRr (4)光颖抗锈×光颖抗锈:ppRr×ppRr 9.大麦的刺芒R对光芒r为显性,黑稃B对白稃b为显性。现有甲品种为白稃,但具有刺芒;而乙品种为光芒,但为黑稃。怎样获得白稃光芒的新品种?(设品种的性状是纯合的) 答:甲、乙两品种的基因型分别为bbRR和BBrr,将两者杂交,得到F1(BbRr),经自交得到F2,从中可分离出白稃光芒(bbrr)的材料,经多代选育可培育出白稃光芒的新品种。 10.小麦的相对性状,毛颖P是光颖p的显性,抗锈R是感锈r的显性,无芒A是有芒a的显性,这三对基因之间不存在基因互作。已知小麦品种杂交亲本的基因型如下,试述F1的表现型。 (1)PPRRAa×ppRraa (2)pprrAa×PpRraa (3)PpRRAa×PpRrAa (4)Pprraa×ppRrAa 答:⑴.F1表现型:毛颖抗锈无芒、毛颖抗锈有芒。 ⑵.F1表现型:毛颖抗锈无芒、毛颖抗锈有芒、毛颖感锈无芒、毛颖感锈有芒、光颖抗锈无芒、光颖抗锈有芒、光颖感锈无芒、光颖感锈有芒。 ⑶.F1表现型:毛颖抗锈无芒、毛颖抗锈有芒、光颖抗锈无芒、光颖抗锈有芒。 ⑷.F1表现型:毛颖抗锈有芒、毛颖抗锈无芒、毛颖感锈无芒、毛颖感锈有芒、光颖感锈无芒、光颖抗锈无芒、光颖抗锈有芒、光颖感锈有芒。 11.光颖、抗锈、无芒(ppRRAA)小麦和毛颖、感锈、有芒(PPrraa)小麦杂交,希望从F3选出毛颖、抗锈、无芒(PPRRAA)的小麦10株,在F2群体中至少应选择表现型为毛颖、抗锈、无芒(P_R_A_)小麦几株? 答:解法一:F1:PpRrAaF2中可以产生毛颖、抗锈、无芒表现型的基因型及其比例:PPRRAA:PpRRAA:PPRrAA:PPRRAa:PpRrAA:PPRrAa:PpRRAa:PpRrAa=1:2:2:2:4:4:4:8按照一般方法则:(1/27)+(6/27)×(1/4)+(12/27)×(1/16)+(8/27)×(1/64)=1/8,则至少选择:10/(1/8)=80(株)。
解法二:可考虑要从F3选出毛颖、抗锈、无芒(PPRRAA)的纯合小麦株系,则需在F2群体中选出纯合基因型(PPRRAA)的植株。 因为F2群体中能产生PPRRAA的概率为1/27,所以在F2群体中至少应选择表现为(P_R_A-_)的小麦植株: 1/27=10X X=10×27=270(株) 12.设有3对独立遗传、彼此没有互作、并且表现完全显性的基因Aa、Bb、Cc,在杂合基因型个体AaBbCc(F1)自交所得的F2群体中,求具有5显性和1隐性基因的个体的频率,以及具有2显性性状和1隐性性状的个体的频率。 答:由于F2基因型比为:27:9:9:9:3:3:3:1而27中A_B_C_中的基因型:AABBCC:AABBCc:AABbCc:AaBBCC:AaBBCc:AaBbCC:AaBbCc (1)5个显性基因,1个隐性基因的频率为: (2)2个显性性状,一个隐性性状的个体的频率: 13.基因型为AaBbCcDd的F1植株自交,设这四对基因都表现为完全显性,试述F2群体中每一类表现型可能出现的频率。在这一群体中,每次任取5株作为一样本,试述3株全部为显性性状、2株全部为隐性性状,以及2株全部为显性性状、3株全部为隐性性状的样本可能出现的频率各为多少? 答:AaBbCcDd:F2中表现型频率:(3/4+1/4)4=81:27:27:27:27:54/4:54/4:54/4:54/4:3:3:3:3:1 ⑴.5株中3株显性性状、2株隐性性状频率为: (81/256)3×(1/256)2=0.0316763×0.0000152587=0.00000048334 ⑵.5株中3株显性性状、3株隐性性状频率为: (81/256)2×(1/256)3=(6561/85536)×(1/16777216)=0.0767045×0.0000000596046=0.00000000457194 14.设玉米子粒有色是独立遗传的三显性基因互作的结果,基因型为A_C_R的子粒有色,其余基因型的子粒均无色。有色子粒植株与以下3个纯合品系分别杂交,获得下列结果: (1)与aaccRR品系杂交,获得50%有色子粒 (2)与aaCCrr品系杂交,获得25%有色子粒 (3)与AAccrr品系杂交,获得50%有色子粒 问这些有色子粒亲本是怎样的基因型? 答:
⑴.基因型为:AACcR_或AaCCR_或AaCcR_或AACcrr或AaCCrr ⑵.基因型为:AaC_Rr或AaccRr ⑶.基因型为:A_CcRR或A_CCRr 15.萝卜块根的形状有长形的、圆形的、有椭圆型的,以下是不同类型杂交的结果: 长形×圆形--595椭圆型 长形×椭圆形--205长形,201椭圆形 椭圆形×圆形--198椭圆形,202圆形 椭圆形×椭圆形--58长形112椭圆形,61圆形 说明萝卜块根属于什么遗传类型,并自定义基因符号,标明上述各杂交亲本及其后裔的基因型? 答:由于后代出现了亲本所不具有的性状,因此属于基因互作中的不完全显性作用。 设长形为aa,圆形为AA,椭圆型为Aa。 (1)aa×AAAa (2)aa×AaAa:aa (3)Aa×AAAA:Aa=198:202=1:1 (4)Aa×AaAA:Aa:aa=61:112:58=1:2:1 16.假定某个二倍体物种含有4个复等位基因(如a1、a2、a3、a4),试决定在下列三种情况下可能有几种基因组合? ⑴.一条染色体;⑵.一个个体;⑶.一个群体。 答:a1、a2、a3、a4为4个复等位基因,故: ⑴.一条染色体上只能有a1或a2或a3或a4; ⑵.一个个体:正常的二倍体物种只含有其中的两个,故一个个体的基因组合是a1a1或a2a2或a3a3或a4a4或a1a2或a1a3或a1a4或a2a3或a2a4或a3a4; ⑶.一个群体中则a1a1、a2a2、a3a3、a4a4、a1a2、a1a3、a1a4、a2a3、a2a4、a3a4等基因组合均可能存在。
第五章本章习题 1.试述交换值、连锁强度和基因之间距离三者的关系。 答:交换值是指同源染色体的非姐妹染色单体间有关基因的染色体片段发生交换的频率,或等于交换型配子占总配子数的百分率。交换值的幅度经常变动在0~50%之间。交换值越接近0%,说明连锁强度越大,两个连锁的非等位基因之间发生交换的孢母细胞数越少。当交换值越接近50%,连锁强度越小,两个连锁的非等位基因之间发生交换的孢母细胞数越多。由于交换值具有相对的稳定性,所以通常以这个数值表示两个基因在同一染色体上的相对距离,或称遗传距离。交换值越大,连锁基因间的距离越远;交换值越小,连锁基因间的距离越近。 2.试述连锁遗传与独立遗传的表现特征及细胞学基础。 答:独立遗传的表现特征:如两对相对性状表现独立遗传且无互作,那么将两对具有相对性状差异的纯合亲本进行杂交,其F1表现其亲本的显性性状,F1自交F2产生四种类型:亲本型:重组型:重组型:亲本型,其比例分别为9:3:3:1。如将F1与双隐性亲本测交,其测交后代的四种类型比例应为1:1:1:1。如为n对独立基因,则F2表现型比例为(3:1)n的展开。独立遗传的细胞学基础是:控制两对或n对性状的两对或n对等位基因分别位于不同的同源染色体上,在减数分裂形成配子时,每对同源染色体上的每一对等位基因发生分离,而位于非同源染色体上的基因之间可以自由组合。连锁遗传的表现特征:如两对相对性状表现不完全连锁,那么将两对具有相对性状差异的纯合亲本进行杂交,其F1表现其亲本的显性性状,F1自交F2产生四种类型:亲本型、重组型、重组型、亲本型,但其比例不符合9:3:3:1,而是亲本型组合的实际数多于该比例的理论数,重组型组合的实际数少于理论数。如将F1与双隐性亲本测交,其测交后代形成的四种配子的比例也不符合1:1:1:1,而是两种亲型配子多,且数目大致相等,两种重组型配子少,且数目也大致相等。连锁遗传的细胞学基础是:控制两对相对性状的两对等位基因位于同一同源染色体上形成两个非等位基因,位于同一同源染色体上的两个非等位基因在减数分裂形成配子的过程中,各对同源染色体中非姐妹染色单体的对应区段间会发生交换,由于发生交换而引起同源染色体非等位基因间的重组,从而打破原有的连锁关系,出现新的重组类型。由于F1植株的小孢母细胞数和大孢母细胞数是大量的,通常是一部分孢母细胞内,一对同源染色体之间的交换发生在某两对连锁基因相连区段内;而另一部分孢母细胞内该两对连锁基因相连区段内不发生交换。由于后者产生的配子全是亲本型的,前者产生的配子一半是亲型,一半是重组型,所以就整个F1植株而言,重组型的配子数就自然少于1:1:1:1的理论数了。 3.大麦中,带壳(N)对裸粒(n)、散穗(L)对密穗(l)为显性。今以带壳、散穗与裸粒、密穗的纯种杂交,F1表现如何?让F1与双隐性纯合体测交,其后代为:带壳、散穗201株,裸粒、散穗18株,带壳、密穗
20株,裸粒、密穗203株。试问,这两对基因是否连锁?交换值是多少?要使F2出现纯合的裸粒散穗20株,至少要种多少株? 答:F1表现为带壳散穗(NnLl)。 F2不符合9:3:3:1的分离比例,亲本组合数目多,而重组类型数目少,所以这两对基因为不完全连锁。 交换值%=((18+20)/(201+18+20+203))×100%=8.6% F1的两种重组配子Nl和nL各为8.6%/2=4.3%,亲本型配子NL和nl各为(1-8.6%)/2=45.7%; 在F2群体中出现纯合类型nnLL基因型的比例为: 4.3%×4.3%=18.49/10000, 因此,根据方程18.49/10000=20/X计算出,X=10817,故要使F2出现纯合的裸粒散穗20株,至少应种10817株。 4.在杂合体ABy//abY内,a和b之间的交换值为6%,b和y之间的交换值为10%。在没有干扰的条件下,这个杂合体自交,能产生几种类型的配子?在符合系数为0.26时,配子的比例如何? 答:这个杂合体自交,能产生ABy、abY、aBy、AbY、ABY、aby、Aby、aBY8种类型的配子。 在符合系数为0.26时,其实际双交换值为:0.26×0.06×0.1×100=0.156%,故其配子的比例为:ABy42.078:abY42.078:aBy2.922:AbY2.922:ABY4.922:aby4.922:Aby0.078:aBY0.078。 5.a和b是连锁基因,交换值为16%,位于另一染色体上的d和e也是连锁基因,交换值为8%。假定ABDE和abde都是纯合体,杂交后的F1又与双隐性亲本测交,其后代的基因型及其比例如何? 答:根据交换值,可推测F1产生的配子比例为(42%AB:8%aB:8%Ab:42%ab)×(46%DE:4%dE:4%De:46%de),故其测交后代基因型及其比例为: AaBbDdEe19.32:aaBbDdEe3.68:AabbDdEe3.68:aabbDdEe19.32: AaBbddDEe1.68:aaBbddEe0.32:AabbddEe0.32:aabbddEe1.68: AaBbDdee1.68:aaBbDdee0.32:AabbDdee0.32:aabbDdee1.68: AaBbddee19.32:aaBbddee3.68:Aabbddee3.68:aabbddee19.32。 6.a、b、c3个基因都位于同一染色体上,让其杂合体与纯隐性亲本测交,得到下列结果:+++74a++106++c382a+c5+b+3ab+364+bc98abc66
试求这3个基因排列的顺序、距离和符合系数。 答:根据上表结果,++c和ab+基因型的数目最多,为亲本型;而+b+和a+c基因型的数目最少,因此为双交换类型,比较二者便可确定这3个基因的顺序,a基因位于中间。 则这三基因之间的交换值或基因间的距离为: ab间单交换值=((3+5+106+98)/1098)×100%=19.3% ac间单交换值=((3+5+74+66)/1098)×100%=13.5% bc间单交换值=13.5%+19.3%=32.8% 其双交换值=(3+5/1098)×100%=0.73% 符合系数=0.0073/(0.193×0.135)=0.28 这3个基因的排列顺序为:bac;ba间遗传距离为19.3%,ac间遗传距离为13.5%,bc间遗传距离为32.8%。 7.已知某生物的两个连锁群如下图,试求杂合体AaBbCc可能产生的类型和比例。 答:根据图示,bc两基因连锁,bc基因间的交换值为7%,而a与bc连锁群独立,因此其可能产生的配子类型和比例为: ABC23.25:Abc1.75:AbC1.75:Abc23.25:aBC23.25:aBc1.75:abC1.75:abc23.25 8.纯合的匍匐、多毛、白花的香豌豆与丛生、光滑、有色花的香豌豆杂交,产生的F1全是匍匐、多毛、有色花。如果F1与丛生、光滑、白花又进行杂交,后代可望获得近于下列的分配,试说明这些结果,求出重组率。 匍、多、有6%丛、多、有19% 匍、多、白19%丛、多、白6% 匍、光、有6%丛、光、有19% 匍、光、白19%丛、光、白6% 答:从上述测交结果看,有8种表型、两类数据,该特征反映出这3个基因有2个位于同一染色体上连锁遗传,而另一个位于不同的染色体上独立遗传。又从数据的分配可见,匍匐与白花连锁,而多毛为独立遗传。匍匐与白花的重组值为24%。假定其基因型为:匍匐AA、多毛BB、白花cc,丛生aa、光滑bb、有色花CC。则组合为: AABBcc×aabbCC ↓ AaBbCc×aabbcc ↓AaBbCc6:AaBbcc19:aaBbCc19:aaBbcc6:AabbCc6:Aabbcc19:aabbCc19:aabbcc6 9.基因a、b、c、d位于果蝇的同一染色体上。经过一系列杂交后得出如下交换值: 基因 交换值
a与c 40% a与d 25% b与d 5% b与c 10% 试描绘出这4个基因的连锁遗传图。 答:其连锁遗传图为: 10.脉孢菌的白化型(al)产生亮色子囊孢子,野生型产生灰色子囊孢子。将白化型与野生型杂交,结果产生: 129个亲型子囊-孢子排列为4亮4灰, 141个交换型子囊----孢子排列为2:2:2:2或2:4:2 问al基因与着丝点之间的交换值是多少? 答:交换值=[141/(141+129)]×100%×1/2=26.1% 11.果蝇的长翅(Vg)对残翅(vg)是显性,该基因位于常染色体上;红眼(W)对白眼(w)是显性,该基因位于X染色体上。现让长翅红眼的杂合体与残翅白眼的纯合体交配,所产生的基因型如何? 答:假如杂合体为双杂合类型,则有两种情况: (1)♀vgvgXwXw×VgvgXWY♂ ↓ VgvgXWXwvgvgXWXwVgvgXwYvgvgXwY (2)♀VgvgXWXw×vgvgXwY♂ ↓ VgvgXWXwVgvgXwXwvgvgXWXwvgvgXwXw VgvgXWYVgvgXwYvgvgXWYvgvgXwY 12.何谓伴性遗传、限性遗传和从性遗传?人类有哪些性状是伴性遗传的? 答:伴性遗传是指性染色体上基因所控制的某些性状总是伴随性别而遗传的现象。 限性遗传是指位于Y染色体(XY型)或W染色体(ZW型)上的基因所控制的遗传性状只局限于雄性或雌性上表现的现象。 从性遗传是指不含于X及Y染色体上基因所控制的性状,而是因为内分泌及其它因素使某些性状或只出现雌方或雄方;或在一方为显性,另一方为隐性的现象。 人类中常见的伴性遗传性状如色盲、A型血友病等。 13.设有两个无角的雌羊和雄羊交配,所生产的雄羊有一半是有角的,但生产的雌羊全是无角的,试写出亲本的基因型,并作出解释。 答:设无角的基因型为AA,有角的为aa,则亲本的基因型为: XAXa(无角)×XAY(无角)
↓ XAXA(无角)XAY(无角) XAXa(无角)XaY(有角) 从上述的基因型和表现型看,此种遗传现象属于伴性遗传,控制角的有无基因位于性染色体上,当有角基因a出现在雄性个体中时,由于Y染色体上不带其等位基因而出现有角性状
第六章染色体变异 本章习题 1.植株是显性AA纯合体,用隐性aa纯合体的花粉给它授粉杂交,在500株F1中,有2株表现为aa。如何证明和解释这个杂交结果? 答:这有可能是显性AA株在进行减数分裂时,有A基因的染色体发生断裂,丢失了具有A基因的染色体片断,与带有a基因的花粉授粉后,F1缺失杂合体植株会表现出a基因性状的假显性现象。可用以下方法加以证明: ⑴.细胞学方法鉴定:①.缺失圈;②.非姐妹染色单体不等长。 ⑵.育性:花粉对缺失敏感,故该植株的花粉常常高度不育。 ⑶.杂交法:用该隐性性状植株与显性纯合株回交,回交植株的自交后代6显性:1隐性。 2.玉米植株是第9染色体的缺失杂合体,同时也是Cc杂合体,糊粉层有色基因C在缺失染色体上,与C等位的无色基因c在正常染色体上。玉米的缺失染色体一般是不能通过花粉而遗传的。在一次以该缺失杂合体植株为父本与正常的cc纯合体为母本的杂交中,10%的杂交子粒是有色的。试解释发生这种现象的原因。 答:这可能是Cc缺失杂合体在产生配子时,带有C基因的缺失染色体与正常的带有c基因的染色体发生了交换,其交换值为10%,从而产生带有10%C基因正常染色体的花粉,它与带有c基因的雌配子授粉后,其杂交子粒是有色的。3.某个体的某一对同源染色体的区段顺序有所不同,一个是12·34567,另一个是12·36547("·"代表着丝粒)。试解释以下三个问题: ⑴.这一对染色体在减数分裂时是怎样联会的? ⑵.倘若在减数分裂时,5与6之间发生一次非姐妹染色单体的交换,图解说明二分体和四分体的染色体结构,并指出产生的孢子的育性。 ⑶.倘若在减数分裂时,着丝粒与3之间和5与6之间各发生一次交换,但两次交换涉及的非姐妹染色单体不同,试图解说明二分体和四分体的染色体结构,并指出产生的孢子的育性。 答:如下图说示。 *为败育孢子。
4.某生物有3个不同的变种,各变种的某染色体的区段顺序分别为:ABCDEFGHIJ、ABCHGFIDEJ、ABCHGFEDIJ。试论述这3个变种的进化关系。 答:这3个变种的进化关系为:以变种ABCDEFGHIJ为基础,通过DEFGH染色体片段的倒位形成ABCHGFEDIJ,然后以通过EDI染色体片段的倒位形成ABCHGFIDEJ。 5.假设某植物的两个种都有4对染色体:以甲种与乙种杂交得F1,问F1植株的各个染色体在减数分裂时是怎样联会的?绘图表示联会形象。 甲种 乙种 ABCDE FGHIJ ADCBE FGMNO ----- ----- ----- ----- ----- ----- ----- ----- ABCDE FGHIJ ADCBE FGMNO KLMNO PQRST KLHIJ PQRST ----- ----- ----- ----- ----- ----- ----- ----- KLMNO PQRST KLHIJ PQRST 答:F1植株的各个染色体在减数分裂时的联会。 6.玉米第6染色体的一个易位点(T)距离黄胚乳基因(Y)较近,T与Y之间的重组率为20%。以黄胚乳的易位纯合体与正常的白胚乳纯系(yy)杂交,试解答以下问题: ⑴.F1和白胚乳纯系分别产生哪些有效配子?图解分析。 ⑵.测交子代(F1)的基因型和表现型(黄粒或白粒,完全不育或半不育)的种类和比例如何?图解说明。
答: 7.曾使叶基边缘有条纹(f)和叶中脉棕色(bm2)的玉米品系(ffbm2bm2),叶基边缘和中脉色都正常的易位纯合体(FFBm2Bm2TT)杂交,F1植株的叶边缘和脉色都正常,但半不育。检查发现该F1的孢母细胞在粗线期有十字形的四分体。使全隐性的纯合亲本与F1测交,测交子代的分离见下表。已知F-f和Bm2-Bm2本来连锁在染色体1的长臂上,问易位点(T)与这两对基因的位置关系如何?叶基边缘有无白条纹中脉色育 性半不育全育36(无)正常99637(有)棕色14038(无)棕色671239(有)正常153 答:⑴.
叶基边缘有无白条纹的比例为1:1:1:1。易位使连锁在同一条染色体上的F-f和Bm2-bm2基因改变为分属于不同的染色体,呈现自由组合规律。因此易位点T在这两基因的中间。 ⑵.易位点T与正常基因之间的遗传距离:F-T为7.16%、Bm2-T为45.52%。 其中:FtBm2和fFbm2为双交换,则: 双交换值=((6+1)/279)=2.51% 单交换值:F-T=((12+1)/279)+2.51%=7.16% Bm2-T=((53+67)/279)+2.51%=45.52%叶基边缘有无白条纹中脉色育 性半不育(T)全育(t)36(F)Bm299637(f)bm214038(F)bm2671239(f)Bm2153 8.某同源四倍体为AaaaBBbb杂合体,A-a所在染色体与B-b所在染色体是非同源的,而且A为a的完全显性,B为b的完全显性。试分析该杂合体的自交子代的表现型比例(设染色体随机分离)。 答: AaaaBBbbF2表现型比例: 自交 配子5AaB-5aaB-1Aabb1aabb5AaB-25AaB-AaB-25AaB-aaB-5AaB-A-bb5AaB-aabb5aaB-25aaB-A-B-25aaB-aaB-5aaB-A-bb5aaB-aabb1Aabb5AabbA-B-5A-abbaaB-1AabbA-bb1Aabbaabb1aabb5aabbA-B-5aabbaaB-1aabbA-bb1aabbaabb
故表型总结为:105A---B---:35aaaaB---:3A---bbbb:1aaaabbbb 9.普通小麦的某一单位性状的遗传常常是由3对独立分配的基因共同决定的,这是什么原因?用小麦属的二倍体种、异源四倍体种和异源六倍体种进行电离辐射处理,哪个种的突变型出现频率最高?哪个最低?为什么? 答:这是因为普通小麦是异源六倍体,其编号相同的三组染色体(如1A1B1D)具有部分同源关系,因此某一单位性状常常由分布在编号相同的三组染色体上的3对独立基因共同决定。如对不同倍数的小麦属进行电离辐射处理,二倍体种出现的突变频率最高,异源六倍体种最低。因为异源六倍体有三组染色体组成,某组染色体某一片段上的基因诱发突变,其编号相同的另二组对应的染色体片段上的基因具有互补作用,可以弥补其辐射带来的损伤。 10.使普通小麦与圆锥小麦杂交,它们的F1植株的体细胞内应有哪几个染色体组和染色体?该F1植株的孢母细胞在减数分裂时,理论上应有多少个二价体和单价体?F2群体内,各个植株的染色体组和染色体数是否还能同F1一样?为什么?是否还会出现与普通小麦的染色体组和染色体数相同的植株? 答:F1植株体细胞内应有AABBD5个染色体组,共35条染色体,减数分裂时理论上应有14II+7I。F2群体内各植株染色体组和染色体数绝大多数不会同F1一样,因为7个单价体分离时是随机的,但也有可能会出现个别与普通小麦的染色体组和染色体数相同的植株。因为产生雌雄配子时,有可能全部7I都分配到一个配子中。 11.马铃薯的2n=48,是个四倍体。曾经获得马铃薯的单倍体,经细胞学的检查,该单倍体在减数分裂时形成12个二价体。据此,你对马铃薯染色体组的组合成分是怎样认识的?为什么? 答:马铃薯是同源四倍体,只有这样,当其是单倍体时,减数分裂才会形成12个二价体。如是异源四倍体话,减数分裂时会形成24个单价体。 12.三体的n+1胚囊的生活力一般远比n+1花粉强。假设某三体植株自交时参与受精的有50%为n+1胚囊,而参与受精的花粉中只有10%是n+1,试分析该三体植株的自交子代群体里,四体所占的百分数、三体所占的百分数和正常2n个体所占的百分数。 答:该三体自交后代的群体为:♀♂90%n10%n+150%n45%2n5%2n+150%n+145%2n+15%2n+2 该三体自交后代的群体里四体(2n+2)、三体(2n+1)、二体(2n)所占的百分数分别为5%、50%、45%。
13.以番茄正常叶型的第6染色体的三体(2n+I6)为母本,以马铃薯叶型(cc)的正常番茄(2n)为父本进行杂交,试问:(1)假设c基因在第6染色体上,使F1群体的三体植株与马铃薯叶型的正常番茄试交,试交子代的染色体数及其表现型(叶型)种类和比例如何?(2)倘若c基因不在第6染色体上,上述试交子代的表现型种类和比例各如何? 答:⑴.假若c基因在第6染色体上,则 (n-1)II+6IIICCC×(n-1)II+6IIcc ↓ (n-1)II+6IIICCc×(n-1)II+6IIcc ↓ 1(n-1)II+6IIICCc+2(n-1)II+6IIICcc+2(n-1)II+6IICc+1(n-1))II+6IIcc 其表现型比例为:正常叶:马铃薯叶=5:1 染色体数比例为:三体:正常=1:1 ⑵.假若c基因不在第6染色体上,则 (n-1)IICC+6III×(n-1)IIcc+6II ↓ (n-1)IICc+6III×(n-1)IIcc+6II ↓ 1(n-1)IICc+6III+1(n-1)IICc+6II+2(n-1)IICc+6III+2(n-1)IICc+6II+1(n-1)IIcc+6III+1(n-1)IIcc+6II+2(n-1)IIcc+6III+2(n-1)IIcc+6II 其后代表现型比例为:正常叶:马铃薯叶=1:1 染色体数比例为:三体:正常=1:1 14.玉米的淀粉质胚乳基因(Su)对甜质胚乳基因(su)为显性。某玉米植株是甜质纯合体(susu),同时是第10染色体的三体(2n+I10)。使该三体植株与粉质纯合的正常玉米(2n)杂交,再使F1群体内的三体植株自交,在F2群体内有1758粒是淀粉质的,586粒是甜质的,问Su-su这对基因是否在第10染色体上?(设染色体随机分离) 答:根据题意,F2群体淀粉质:甜质=1758:586=3:1,可推知这对基因不在第10染色体上。解释: (n-1)IIsusu+10III×(n-1)IISuSu+10II ↓ (n-1)IISusu+10III ↓自交(n-1)ISu+10I(n-1)ISu+10II(n-1)Isu+10I(n-1)Isu+10II(n-1)ISu+10I 淀粉质:甜质=3:1(n-1)ISu+10II(n-1)Isu+10I(n-1)Isu+10II 如在第10染色体上,则 (n-1)II+10IIIsususu×(n-1)II+
10IISuSu ↓ (n-1)II+10IIISususu ↓自交1(n-1)I+10ISu1(n-1)I+10IIsusu2(n-1)I+10Isu2(n-1)I+10IISusu1(n-1)I+10ISu 淀粉质:甜质=27:9=3:11(n-1)I+10IIsusu2(n-1)I+10Isu2(n-1)I+10IISusu 上述是假定三体10IIISususu的分离中n+1和n以同等的比例授精,但实际上三体n+1的配子参与受精的要少于n配子,n+1的花粉更少,因此不可能达到刚好是3:1的比例。因此不在第10染色体上。 15.一般都认为烟草是两个野生种Nicotianasylvestris(2n=24=12II=2X=SS)和N.tomentosiformis(2n=24=12II=2X=TT)合并起来的异源四倍体(2n=48=24II=SSTT)。某烟草单体(2n-1=47)与N.sylvestris杂交的F1群体内,一些植株有36个染色体,另一些植株有35个染色体。细胞学的检查表明,35个染色体的F1植株在减数分裂时联会成11个二价体和13个单价体,试问:该单体所缺的那个染色体属于S染色体组,还是属于T染色体组?如果所缺的那个染色体属于你所解答的那个染色体组的另一个染色体组,上述的35个染色体的F1植株在减数分裂时应该联会成几个二价体和单价体? 答:(1)该单体所缺的那个染色体属于S染色体组,因为具有35个染色体的F1植株在减数分裂时形成了11二价体和13个单价体。 (2)假若该单体所缺的那个染色体属于T染色体组,则35个染色体的F1植株在减数分裂时会形成12二价体和11个单价体。 16.白肋型烟草的茎叶都是乳黄绿色,基因型是yb1yb1yb2yb2隐性纯合体。某植株的基因型内只要有yb1的显性等位基因Yb1或yb2的显性等位基因Yb2中之一个即为正常绿色。曾使白肋型烟草与9个不同染色体(从M到U)的单体杂交,得9个杂交组合的F1,再使白肋型烟草分别回交9个F1群体内的单体植株,得到下表所列的回交子一代。试问Yb1-yb1或Yb2-yb2在哪条染色体上?为什么?F1单体的单体染色体回交子一代的表现种类和株数绿株白肋株M369N288O1917P339Q3212R2712S274T288
U378 答:Yb1-yb1或Yb2-yb2位于O染色体上。下面以Yb2-yb2是否位于O染色体上作相关解释: ⑴.如果Yb2在O染色单体上, (n-2)II+IIyb1yb1+M-UIIyb2yb2×(n-2)II+IIYb1Yb1+OIYb2 ↓ (n-2)II+IIYb1yb1+OIIYb2yb2; (n-2)II+IIYb1yb1+OIyb2×(n-2)II+IIyb1yb1+OIIyb2yb2 ↓(n-2)II+IIYb1yb1+OIyb21绿(n-2)II+IIYb1yb1+OIIyb2yb21绿(n-2)II+IIyb1yb1+OIIyb2yb21白(n-2)II+IIyb1yb1+OIyb21白 则正常株和白肋株的比例为1:1,而上表中只有O染色体单体后代表现为19:17接近于理论比例1:1,故推测Yb2基因位于O染色体上。 同理如Yb1基因位于单体染色体上,也表现为相同的遗传规律,因此Yb1基因也可位于O染色体上。 ⑵.如果不在O染色单体上,则 (n-2)II+IIyb1yb1+IIyb2yb2×(n-3)II+IIYb1Yb1+IIYb2Yb2+OI ↓ (n-2)II+IIYb1yb1+IIYb2yb2; (n-3)II+IIYb1yb1+IIYb2yb2+OI×(n-2)II+IIyb1yb1+IIyb2yb2 ↓((n-3)II+OI)(n-2)II+IIYb1yb1+IIYb2yb21绿株((n-3)II+OI)(n-2)II+IIYb1yb1+IIyb2yb21绿株((n-3)II+OI)(n-2)II+IIyb1yb1+IIYb2yb21绿株((n-3)II+OI)(n-2)II+IIyb1yb1+IIyb2yb21白肋株 则正常株和白肋株的比例为3:1,而上表中只有除O染色体株之外其它染色体单体后代的表现接近于这一理论比例
第七章细菌和病毒的遗传 本章习题 1.解释下列名词:F-菌株、F+菌株、Hfr菌株、F因子、F"因子、烈性噬菌体、温和性噬菌体、溶原性细菌、部分二倍体。 F-菌株:未携带F因子的大肠杆菌菌株。 F+菌株:包含一个游离状态F因子的大肠杆菌菌株。 Hfr菌株:包含一个整合到大肠杆菌染色体组内的F因子的菌株。 F因子:大肠杆菌中的一种附加体,控制大肠杆菌接合过程而使其成为供体菌的一种致育因子。 F"因子:整合在宿主细菌染色体上的F因子,在环出时不够准确而携带有染色体一些基因的一种致育因子。 烈性噬菌体:侵染宿主细胞后,进入裂解途径,破坏宿主细胞原有遗传物质,合成大量的自身遗传物质和蛋白质并组装成子噬菌体,最后使宿主裂解的一类噬菌体。 温和性噬菌体:侵染宿主细胞后,并不裂解宿主细胞,而是走溶原性生活周期的一类噬菌体。 溶原性细菌:含有温和噬菌体的遗传物质而又找不到噬菌体形态上可见的噬菌体粒子的宿主细菌。 部分二倍体:当F+和Hfr的细菌染色体进入F-后,在一个短时期内,F-细胞中对某些位点来说总有一段二倍体的DNA状态的细菌。 2.为什么说细菌和病毒是研究遗传学的好材料? 答:与其他生物体相比,细菌和病毒能成为研究遗传学的好材料,具有以下7个方面的优越性: (1)世代周期短:每个世代以min或h计算,繁殖速度快,大大缩短了实验周期。 (2)易于管理和进行化学分析个体小,繁殖方便,可以大量节省人力、物力和财力;且代谢旺盛,繁殖又快,累积大量的代谢产物。 (3)便于研究基因的突变细菌和病毒均属于单倍体,所有突变都能立即表现出来,不存在显性掩盖隐性的问题。 (4)便于研究基因的作用通过基本培养基和选择培养基的影印培养,很容易筛选出营养缺陷型,利于生化[研究。 (5)便于基因重组的研究通过细菌的转化、转导和接合作用,在一支试管中可以产生遗传性状不相同的后代。 (6)便于用于研究基因结构、功能及调控机制的材料细菌和病毒的遗传物质简单,基因定位和结构分析等易于进行且可用生理生化方法进行基因的表达和调控分析。 (7)便于进行遗传操作细菌质粒和病毒作为载体,已成为高等生物的分子遗传学研究和生物工程的重要工具。 3.试比较大肠杆菌和玉米的染色体组。 答:
大肠杆菌属于原核生物、而玉米是真核生物,二者基因组存在很大的区别: ⑴.基因组大小不同:大肠杆菌DNA以单个染色体的形式存在,长约1100μm,分子量约为2.6×109;玉米以10对染色体存在(n=10),基因组非常庞大。 ⑵.染色体组成不同:大肠杆菌DNA不与组蛋白结合,也不形成核小体结构,是一个封闭的大环结构;而玉米DNA与组蛋白结合,形成典型的核小体结构,呈直线排列,并多级折叠成光学显微镜下可见的染色体结构。 ⑶.大肠杆菌的基因发生突变,在当代个体中即可表现出来,而在玉米中基因组中则存在基因的显隐性关系。 ⑷.DNA合成时期不同:大肠杆菌DNA在整个细胞生长过程中都可进行,而玉米DNA只在细胞周期的S期合成。 ⑸.复制起点不同:大肠杆菌只有一个复制起点,在而玉米存在多个复制起点。 ⑹.DND组成不同:大肠杆菌中一般由单一序列组成,且基因的排列方式非常紧凑,存在重叠基因现象;而玉米中则存在大量的重复序列,许多基因以基因家族方式存在。 4.对两个基因的噬菌体杂交所测定的重组频率如下: a-b+×a+b-3.0% a-c+×a+c-2.0% b-c+×b+c-1.5% 试问:(1)a、b、c3个突变在连锁图上的次序如何?为什么它们之间的距离不是累加的? (2)假定三因子杂交,ab+c×a+bc+,你预期哪种类型的重组体频率最低? (3)计算从⑵所假定的三因子杂交中出现的各种重组类型的频率。 答:⑴.a、b、c3个突变在连锁图上的次序为右图,由于噬菌体的DNA是环状结构,而不是线状排列,因此它们之间的距离不是累加的。 ⑵.根据⑴的三个基因间的连锁距离可知,基因间重组率较低的是ac和bc,因此ab+c+和a+bc两种类型的重组体频率最低。
⑶.根据⑴的重组率可知:c基因在中间: bc间单交换产生acb和a+c+b+的频率共为1.5%; ac间单交换产生a+cb+和ac+b的频率共为2.0%; 双交换ac+b+和a+cb的频率共为0.03%。 5.噬菌体三基因杂交产生以下种类和数目的后代:+++235 pqr270pq+62 p++7+q+40 p+r48+qr4 ++r60 共:726 试问:(1)这一杂交中亲本噬菌体的基因型是什么? (2)基因次序如何? (3)基因之间的图距如何? 答:(1)这一杂交中亲本基因型是+++和pqr; (2)根据杂交后代中双交换类型和亲本基因型,便可推断出基因次序为:qpr或rpq; (3)基因之间的图距:类型基因型数目 比例(%)重组率(%)亲本类型+++235505 pqr270 单交换型Ipq+6212216.8√ √ ++r60 单交换型IIp+r488812.1 √√ +q+40 双交换型p++7111.5√√ +qr4共: 726 18.313.629 pr之间的遗传距离为18.3遗传单位;pq之间的遗传距离为13.6遗传单位;因为有双交换的存在,qr之间的遗传距离为:28.9+2×1.5=31.9遗传单位。 6.试比较转化、接合、转导、性导在细菌遗传物质传递上的异同。 答:
这四种现象的相同之处是:都是细菌的遗传物质DNA在不同的细菌细胞之间传递,从而使受体细胞遗传物质发生重组。 不同之处是:转化是裸露的DNA直接与处于感受态的细胞之间的互作,进入受体细胞,发生重组;接合是由于F因子的整合产生Hfr菌株,在F因子进行转移时,供体菌遗传物质也被带入受体菌,实现重组;性导是Hfr菌株中F因子的错误环出,产生了携带有供体菌遗传物质的F"因子,接合时随F"因子的转移而使供体菌遗传物质导入到受体菌中;转导是细菌的一段染色体被错误地包装在噬菌体的蛋白质外壳内,并通过感染而转移到另一个受体菌内。 7.7.假定你证明对过去一个从未描述过的细菌种有遗传重组,如使ab+菌株与a+b菌株混合培养,形成a+b+、ab的重组类型,试说明将采用哪种方式来确定这种重组是转化、转导还是接合的结果。 答:参照戴维斯的U型管试验,将两菌株放入培养,后代中发现如无重组类型,则该遗传重组类型为接合产生的;后代中如有重组类型,可能是转化或转导产生的;可进一步试验,在U型管中加入DNA酶,检测后代有无重组,如无重组则为该类型为转化产生的,如有则是转导产生的。 8.在接合实验中,Hfr菌株应带有一个敏感的位点(如azis或strs),这样,在发生接合后可用选择培养基消除Hfr供体。试问这个位点距离Hfr染色体的转移起点(O)应该远还是近,为什么? 答:这个位点距离Hfr染色体的转移起点(O)应该是远。 因为如这个敏感位点距转移起点(O)近情况下,Hfr菌株的基因从原点处开始进入受体菌,使得敏感位点较早地重组进受体菌中,在中断杂交后,除去Hfr菌株的同时也除去了重组有敏感位点的重组个体,这样就无法检测敏感位点之后的基因重组距离了。 9.供体菌株为Hfrarg-leu+aziSstrS,受体菌株F-arg+leu-aziRstrS。为检出和收集重组体F-arg+leu+aziR,应用下列哪一种培养基可以完成这一任务,为什么其它的培养基不可以?⑴.基本培养基加链霉素,⑵.基本培养基加叠氮化钠和亮氨酸,⑶.基本培养基加叠氮化钠,⑷.选择培养基中不加精氨酸和亮氨酸,加链霉素,⑸.基本培养基加链霉素和叠氮化钠。 答:3号培养基合适,因为:1号培养基,所有菌株均为链霉素敏感,在该培养基中将抑制所有的菌株;2号培养基,无法区分重组体和受体菌;3号培养基,加叠氮化钠可以抑制供体菌的生长,同时又不加亮氨酸,受体菌也无法生长;4号培养基中,加链霉素将抑制所有菌株;5号培养基,加链霉素也将将抑制所有菌。 10.大肠杆菌3个Hfr菌株利用中断交配技术,分别与营养缺陷型F-菌株交配,获得下表结果:供体位点进入时间(min)HfrP4XHfrKL98HfrRa-2gal+116770thr+945087xyl+73298
lac+25879his+389443ilu+77334arg+621819 试利用上述资料建立一个大肠杆菌染色体图,包括以min表示的图距。并标出各Hfr菌株F因子的插入位点及转移方向。 答:根据上表结果可知各基因位点在不同菌株中的排列顺序:菌株供体位点HfrP4Xlac+gal+his+arg+xyl+ilu+thr+HfrKL98arg+xyl+ilu+thr+lac+gal+his+HfrRa-2ilu+xyl+arg+his+gal+lac+thr+ 11.利用大肠杆菌菌株杂交,一个是a+b+c-d+,另一个是a-b-c+d-。从重组体中选择b+c+基因,而不选a及d的等位基因,当检查b+c+时,大部分都是a-d-。试问:⑴.哪一个菌株是供体?⑵.从这个实验可以得到什么结论? 答:(1)一般重组类型占比例比亲本类型少,当检查b+c+时,大部分都是a-d-,因此a-b-c+d-基因型为受体菌。 (2)本实验说明了F因子插入位点位于bc之前,而离ad较远。 12.如果把一个大肠杆菌放在含λ的培养基上它并不裂解,你是否认为这个大肠杆菌是溶原性的?
答:这个原始大肠杆菌是溶原性的。 因为:当溶原性的大肠杆菌放入含λ噬菌体的培养基中时,由于大肠杆菌本身存在抗超数感染性质,因此不裂解;另外λ噬菌体是温和性噬菌体,侵染大肠杆菌之后,进入溶原状态,并不马上走裂解途径。 13.Hfrmet+thi+pur+×F-met-thi-pur-杂交。中断杂交试验表明,met+最后进入受体。所以只在含thi和pur的培养基上选择met+接合后重组体。检验这些接合后体存在的thi+和pur+,发现各基因型个体数如下: met+thi+pur+280met+thi+pur-0 met+thi-pur+6met+thi-pur-52 试问:(1)选择培养基中为什么不考虑met? (2)基因次序是什么? (3)重组单位的图距有多大? (4)这里为什么不出现基因型met+thi+pur-的个体? 答:(1)因为met+最后进入受体,易于检测出。 (2)基因次序是thi+pur+met+。 (3)重组单位的图距是: (4)在三个位点间发生双交换才有可能发生met+thi+pur-的个体,由于中断杂交的时间短或者所筛选的群体小,未能发现该个体。 14.大肠杆菌中3个位点ara、leu和ilvH是在1/2min的图距内,为了确定三者之间的正确顺序及图距,用转导噬菌体P1侵染原养型菌株ara+leu+ilvH+,然后使裂解物侵染营养缺陷型菌株ara-leu-ilvH-,对每个有选择标记基因进行实验,确定其未选择标记基因的频率,获下表结果:实验选择的标记基因未选择的标记基因1ara+60%leu+1%ilvH+2ilvH+5%ara+0%leu+3ara+ilvH+0%leu+ 根据上表3个实验结果,试说明: (1)三个基因间的连锁顺序如何?(2)这个转导片段的大小。 答:⑴.
三个基因间的连锁顺序:由实验1可知,ara基因距leu基因近,而距ilvH基因远;由实验2可知,ilvH基因距ara基因近,而距leu基因远;由实验3进一步验证,ilvH基因与ara基因间,无leu基因。因此三个基因的连锁顺序为: ⑵.这个转导片段的大小:ilvH基因与ara基因间的并发转导中有1~5%,与leu基因间未发生过转导,因此,这个转导片段的大小是从ilvH位点到ara和leu位点之间。 15.肺炎双球菌中基因型为strSmtl-(mtl+为发酵甘露醇(mannitol)的基因,mtl-不能发酵甘露醇)的细菌在一个试验中由具有strRmtl+的DNA进行转化,在另一个试验中由具有strRmtl-的以及具有strSmtl+的两种DNA混合物进行转化,其结果如下:供体DNA转化产生的基因型的百分数strRmtl-strSmtl+strRmtl+strRmtl-4.30.400.17StrRmtl-+strSmtl-2.80.850.0066 试问: (1)上表中第一横行所列结果说明了什么?为什么? (2)上表中第二横行所列结果说明了什么?为什么? 答:⑴.strRmtl+的比例很小,说明这两个位点的相距较远。因为,DNA转化只能以小片段的形式进入受体,距离远的两个基因同时位于同一个片段的机会小,并发转化的机会也小。 ⑵.两基因位于不同的片段上,并发转化的概率是两个位点单个转化的概率的乘积,因此产生strRmtl+基因型的个体更少,且明显少于共存于同一染色体上的两个位点的共转化。 16.在普遍性转导中,供体大肠杆菌细胞的基因型是trpC+pyrF-trpA-,受体细胞的基因型是trpC-pyrF+trpA+。由P1噬菌体媒介转导,对trpC+进行选择,用选择的细胞进一步检查其它基因的转导情况,得到以下的结果:基因型后代数目trpC+pyrF-trpA-274trpC+pyrF+trpA-279trpC+pyrF-trpA+2trpC+pyrF+trpA+46试问: (1)这3个基因的次序是什么? (2)TrpC和pyrF以及trpC和trpA的合转导频率是多少? (3)假定P1染色体长为10mm,这些基因之间的物理距离是多少? 答:⑴.这3个基因的次序: 由上表可知,后代数目最少的基因型为trpC+pyrF-trpA+,因为三个基因位点中,只有发生了双交换的频率是最少的,可以推断基因顺序为:trpCtrpApyrF; ⑵.
TrpC和pyrF以及trpC和trpA的合转导频率: TrpC和pyrF的合转导频率是: trpC和trpA的合转导频率是: ⑶.假定P1染色体长为10mm,这些基因之间的物理距离: TrpC和pyrF的物理距离是: trpC和trpA的物理距离是: 两个基因同在一起转导就是合转导,合转导的频率愈高,表明两个基因在染色体上的距离愈近,连锁愈密切,测定两基因的合转导频率就可以确定基因之间的次序和距离。通过观察两个基因的转导,即两因子转导(two-factortransduction),计算并比较每两个基因之间的合转导频率,就可以确定三个基因或三个以上基因在染色体上的排列顺序。 17.大肠杆菌Hfrgal+lac+(A)与F-gal-lac-(B)杂交,A向B转移gal+比较早而且频率高,但是转移lac+迟而且效率低。菌株B的gal+重组体仍旧是F-。从菌株A可以分离出一个变体叫做菌株C,菌株C向B转移lac+早而且频率高,但不转移gal+。在C×B的杂交中,B菌株的lac+重组体一般是F+。问菌株C的性质是什么? 答:根据题意可推断,Hfr菌株A是高频同源重组菌株,F因子插入的位置是位于gal+lac+之间,且gal+基因靠近于F因子转移的起点,lac+基因则相反,因此A向转移gal+比较早而且频率高,但是转移lac+迟而且效率低。由于细菌染色体很长,一般容易中断,很难转移完整的一个F因子,因此,菌株B的gal+重组体仍为F-。 从菌株A的变体菌株C,可推断为,由于lac+基因靠近F因子的另一端,F因子环出时错误地包装了lac+基因而未包裹gal+基因,因此,C×B的杂交中,B菌株的lac+重组体一般是F+。菌株C向B转移lac+早,而且频率高,但不转移gal+。 18.在大肠杆菌中发现了一个带有麦芽糖酶基因(mal)的F"因子。将F"mal+引入F-mal-菌株。这样所产生的细胞多数能转移F"mal+到F-细胞中,偶然有些细胞可以从mal-基因开始把整个细菌染色体转移到F-中。这些细胞可分为两类:(a)转移mal+很早,mal-很迟;(b)转移mal-很早,mal+很迟。 画出F"因子与染色体相互作用的图,表明(a)型细胞最大的可能是如何产生的,(b)型细胞又是如何产生的。 答:
有些细胞可以从mal-基因开始把整个细菌染色体转移到F-中,那么这些细胞肯定是Hfr类型,假定F"因子带有A、B、C、D四个区域,转移切口(O)发生在C和D之间,mal+基因位于A与B之间,则(a)型细胞产生最大的可能是:F"mal+整合在细胞染色体上,位置恰好相邻于mal-基因,而mal+基因紧挨着转移切口位点,而mal-基因则相反,具体见下图。 (b)型细胞产生最大的可能是:F"mal+先与细胞染色体上的mal-基因发生同源重组,产生F"mal-因子,该因子再与主染色体发生整合,位置同(a),具体过程见下图。
第八章基因表达与调控 本章习题 1.经典遗传学和分子遗传学关于基因的概念有何不同? 答:孟德尔把控制性状的因子称为遗传因子;约翰生提出基因(gene)这个名词,取代遗传因子;摩尔根等对果蝇、玉米等的大量遗传研究,建立了以基因和染色体为主体的经典遗传学。 经典遗传学认为:基因是一个最小的单位,不能分割;既是结构单位,又是功能单位。具体指:⑴.基因是化学实体:以念珠状直线排列在染色体上;⑵.交换单位:基因间能进行重组,而且是交换的最小单位。⑶.突变单位:一个基因能突变为另一个基因。⑷.功能单位:控制有机体的性状。 分子遗传学认为:⑴.将基因概念落实到具体的物质上,并给予具体内容:一个基因是DNA分子上的一定区段,携带有特殊的遗传信息。⑵.基因不是最小遗传单位,而是更复杂的遗传和变异单位:例如在一个基因区域内,仍然可以划分出若干起作用的小单位。现代遗传学上认为:①.突变子:是在性状突变时,产生突变的最小单位。一个突变子可以小到只有一个碱基对,如移码突变。②.重组子:在性状重组时,可交换的最小单位称为重组子。一个交换子只包含一个碱基对。③.顺反子:表示一个作用的单位,基本上符合通常所描的基因大小或略小,包括的一段DNA与一个多链的合成相对应,即保留了基因是功艿ノ坏慕馐汀"?分子遗传学对基因概念的新发展:结构基因:指可编码RNA或蛋白质的一段DNA序列。调控基因:指其表达产物参与调控其它基因表达的基因。重叠基因:指在同一段DNA顺序上,由于阅读框架不同或终止早晚不同,同时编码两个以上基因的现象。隔裂基因:指一个基因内部被一个或更多不翻译的编码顺序即内含子所隔裂。跳跃基因:即转座因子,指染色体组上可以转移的基因。假基因:同已知的基因相似,处于不同的位点,因缺失或突变而不能转录或翻译,是没有功能的基因。 2.有一个双突变杂合二倍体,其基因型是+a//b+,如果有互补作用表示什么?如果无互补作用,表示什么? 答:有互补作用:表示该表现型为野生型,a、b两突变不是等位的,是代表两个不同的基因位点。 无互补作用:表示该表现型为突变型,a、b两突变是等位的,是代表同一个基因位点的两个基因座。 3.用T4噬菌体一个特殊基因区的不同突变体研究互补作用,获得下列资料。试根据这个资料判断,本区应有几个顺反子? 1234561-+-+--2+-+-++3-+-+--4+-+-++5-+-+--
6-+-+-- +为互补,-为无互补; 答:从第一行可以看出,突变体1与突变体3、5、6不互补,隶属于同一个基因位点,即为同一个顺反子;而与突变体2、4不互补,不在同一个基因位点上;第三、五、六行则进一步验证了第一行的推断。 从第二、四行可以看出,突变体2、4不互补,隶属于同一个基因位点,即为同一个顺反子;而与突变体1、3、5、6互补,不在同一个基因位点上,即分属于不同的顺反子。结论: 本区共有两个顺反子,突变体1、3、5、6同属于一个顺反子;突变体2、4同属于一个顺反子。 4.举例说明基因的微细结构是如何建立的。 答:以本泽尔利用经典的噬菌体突变和重组技术,对T4噬菌体rII区基因微细结构的分析为例。 原理:r+野生型T4噬菌体:侵染E.coliB株和K12株,形成的噬菌斑小而模糊;rII突变型T4噬菌体:只能侵染B株,不能侵染K12(λ)株,形成的噬菌斑大而清楚。 利用上述特点,让两个rII突变型杂交,接种K12(λ)株选择重组体r+,计算出两个r+突变座位间的重组频率,具体过程见右图。 重组值计算: rxry的数量与r+r+相同,计算时r+r+噬菌体数×2。可以获得小到0.001%,即十万分之一的重组值。利用大量rⅡ区内二点杂交的结果,绘制出rⅡ区座位间微细的遗传图: 5.举例说明基因是如何控制遗传性状表达的。 答:
基因对于遗传性状表达的作用可分为直接与间接两种形式。 (1)如果基因的最后产品是结构蛋白或功能蛋白,基因的变异可以直接影响到蛋白质的特性,从而表现出不同的遗传性状。例如人的镰形红血球贫血症。红血球碟形HbA型产生两种突变体Hbs、Hbc红血球镰刀形。血红蛋白分子有四条多肽链:两条α链(141个氨基酸/条)、两条β链(146个氨基酸/条)。HbA、Hbs、Hbc氨基酸组成的差异在于β链上第6位上氨基酸,HbA第6位为谷氨酸(GAA)、Hbs第6位为缬氨酸(GUA)、Hbc第6位为赖氨酸(AAA)。 基因突变会最终影响到性状改变,产生贫血症的原因:仅由单个碱基的突变,引起氨基酸的改变,导致蛋白质性质发生变化,直接产生性状变化。由正常的碟形红血球转变为镰刀形红血球,缺氧时表现贫血症。 (2)更多的情况下,基因是通过酶的合成,间接影响生物性状的表达,例如豌豆:圆粒(RR)×皱粒(rr)产生F1圆粒(Rr),自交产生F2,1/4表现为皱粒(rr)。rr的表现型为皱粒,是因为缺少一种淀粉分支酶(SBE)所致。SBE控制淀粉分支点的形成,rr豌豆的SBE不正常,带有一段0.8kb的插入片段,结果形成异常mRNA,不能形成淀粉分支酶。在种子发育过程中,不能合成淀粉导致积累蔗糖和大量的水分。随着种子的成熟,皱粒基因型(rr)种子比圆粒基因型种子失水快,结果形成皱粒种子表现型。而F1圆粒(Rr)杂合体中,有一个正常的R基因,可以产生SBE酶,能够合成淀粉,表现为圆粒。本例说明R与r基因控制豌豆子粒的性状不是直接的,而是通过指导淀粉分支酶的合成间接实现的。 6.试说明正调控与负调控的区别。 答:转录水平的调控通常可归为正调控与负调控两种。正调控与负调控并非互相排斥的两种机制,而是生物体适应环境的需要,有的系统既有正调控又有负调控。 正调控是经诱导物诱导转录的调控机制。诱导物通常与蛋白质结合,形成一种激活子复合物,与基因启动子DNA序列结合,激活基因起始转录,使基因处于表达的状态;负调控是细胞中阻遏物阻止基因转录过程的调控机制。阻遏物与DNA分子的结合,阻碍RNA聚合酶转录,使基因处于关闭状态。 真核生物以正调控为主;原核生物以负调控为主。 降解代谢途径中既有正调控又有负调控;合成代谢途径中通常以负调控来控制产物自身的合成。 7.假设R基因编码的蛋白质,是S基因转录的负调控子。试问⑴.在R-突变体中S基因是否转录?⑵.如果R基因产物是S基因转录的正调控子,结果又有什么不同? 答:(1)在R-突变体中S基因是否转录有两种情况:如果S基因转录属负调控系统,则在R-突变体中,S基因转录;如果S基因转录既有负调控又有正调控来共同控制,则该突变体中,尽管基因失活,如无正调控开启,仍无法转录基因。 (2)如果R基因产物是S基因转录的正调控子,则在该突变体中,R基因失活,则无正常的正调控因子,转录系统不开启,基因S不转录。 8.试述乳糖操纵元模型。 答:1961年,JacobF.和Monod
J.的乳糖操纵元模型:乳糖操纵元阐述的是一个基因簇内结构基因及其调控位点的表达调控方式。包括编码乳糖代谢酶的3个结构基因及其邻近的调控位点,即一个启动子和1个操纵子,还有位于上游的抑制基因。大肠杆菌乳糖代谢的调控需要三种酶参加:①.β-半乳糖酶由结构基因lacZ编码,将乳糖分解成半乳糖和葡萄糖;②.渗透酶由结构基因lacY编码,增加糖的渗透,易于摄取乳糖和半乳糖;③.转乙酰酶由结构基因lacA编码,β-半乳糖转变成乙酰半乳糖。三个结构基因受控于同一个调控系统,大量乳糖时,大肠杆菌三种酶的数量急剧增加,几分钟内达到千倍以上,这三种酶能够成比例地增加;乳糖用完时,这三种酶的合成也即同时停止。 在乳糖操纵元中,lacI基因编码一种阻遏蛋白,该蛋白至少有两个结合位点,一个与DNA结合,另一个与乳糖结合。当没有乳糖时,lacI基因产生的阻遏蛋白,结合在操纵子位点的DNA序列上,阻止RNA聚合酶起始转录结构基因。在有乳糖时,乳糖与阻遏蛋白结合,使其空间构型发生改变,而不能与操纵子DNA结合,这样RNA聚合酶起始转录结构基因,产生乳糖代谢酶,开始代谢乳糖。因此乳糖操纵元是一种负调控机制。 9.指出下列每一种部分二倍体①.是否合成β-半乳糖苷酶,②.是诱导型还是组成型?(斜线左侧是质粒基因型,右侧是染色体基因型) (1)lacZ+lacY-/lacZ-lacY+ (2)lacOClacZ-lacY+/lacZ+lacy- (3)lacP-lacZ+/lacOClacZ- (4)lacI+lacP-lacZ+/lacI-lacZ+ 答:⑴.lacZ+lacY-/lacZ-lacY+质粒DNA能合成β-半乳糖苷酶,是诱导型;⑵.lacOClacZ-lacY+/lacZ+lacY-染色体DNA能合成β-半乳糖苷酶,是诱导型;⑶.lacP-lacZ+/lacOClacZ-均不能合成β-半乳糖苷酶,是组成型;⑷.lacI+lacP-lacZ+/lacI-lacZ+染色体DNA能合成β-半乳糖苷酶,是组成型。 10.试述色氨酸操纵元和阿拉伯糖操纵元模型。 答:色氨酸操纵元模型 由JacobF.和MonodJ.提出,是具有合成代谢途径典型的操纵元模型。色氨酸操纵元模型结构,5种结构基因trpE,D,C,B,A编码色氨酸合成有关的5种酶;调控结构:启动子、操纵子、前导序列、弱化子;阻遏物trpR基因:与trp操纵元相距较远。大量色氨酸时,大肠杆菌5种酶的转录同时受到抑制;色氨酸不足时,这5种酶的基因开始转转录。色氨酸:作为阻遏物而不是诱导物参与调控结构基因的转录,因此,trp操纵元是一个典型的可阻遏的操纵元模型(repressibleoperon)。包括有两类调控机理: (1).阻遏调控 trpR基因编码无辅基阻遏物,与色氨酸结合,产生有活性的色氨酸阻遏物,与操纵子结合,阻止转录;色氨酸不足,阻遏物三维空间结构发生变化,不能与操纵子结合,操纵元开始转录;色氨酸浓度升高:色氨酸与阻遏物结合,空间结构发生变化,可与操纵子结合,阻止转录; (2).弱化子调控 前导序列可翻译出一段14个氨基酸的短肽,在该短肽的第10,11位置上是两个色氨酸的密码子;两个密码子之后是一段mRNA序列,该序列可分为四个区段,区段间可互补配对,形成不同的二级结构。原核生物具有边转录边翻译的特点,前导序列中,核糖体位置将决定形成哪种二级结构,从而决定弱化子是否可形成终止信号。①.当有色氨酸时,可完整地翻译出短肽,核糖体停留在终止密码子处,邻近区段2位置,阻碍了2,3配对,使3,4区段配对形成发夹结构终止子,RNA酶在弱化子处终止,不能向前移动。②.
如缺乏色氨酸,核糖体到达色氨酸密码子时,由于没有色氨酰tRNA的供应,停留在氨酸密码子位置,位于区段1,使区段2与区段3配对,区段4无对应序列配对呈单链状态,RNA聚合酶顺利弱化子,继续向前移动,转录出完整的多顺反子序列。 阿拉伯糖操纵元模型 阿拉伯糖操纵元是控制分解代谢途径的另一调控系统。其特点是调节蛋白既可以起正调控作用,又可以起负调控作用。 组成结构包括3个结构基因B、A、D和三个调控位点R、O、I,其中R是araC基因编码调节蛋白AraC蛋白,O包括两部分,O1不参与调控、O2是AraC蛋白负调控结合位点,I是调节位点,CAP-cAMP复合物结合位点,AraC蛋白正调控结合位点。 调控调控机理:诱导物阿拉伯糖和cAMP同时存在,阿拉伯糖与araC蛋白复合物结合在I位点,CAP-cAMP复合物结合I位点,基因转录开启。在没有诱导物阿拉伯糖和cAMP时,AraC蛋白同时与I和O2结合,DNA构型发生改变,形成一个紧密的环结构,抑制表达。 11.举例说明阻遏物与无辅基阻遏物的区别。 答:阻遏物一般出现在代谢调控途径中,而无辅基阻遏物则出现合成途径中的调控。两者具有明显不同的调控机理: 阻遏物:如在乳糖操纵子模型中的阻遏蛋白,由lacI基因编码,该蛋白至少有两个结合位点,一个与DNA结合、另一个与乳糖结合。当没有乳糖时,lacI基因产生的阻遏蛋白,结合在操纵子位点的DNA序列上,阻止RNA聚合酶起始转录结构基因。在有乳糖时,乳糖与阻遏蛋白结合,使其空间构型发生改变,而不能与操纵子DNA结合,这样RNA聚合酶起始转录结构基因,产生乳糖代谢酶,开始代谢乳糖。 无辅基阻遏物:如在色氨酸操纵元模型中,trpR基因编码无辅基阻遏物,与色氨酸结合,产生有活性的色氨酸阻遏物,与操纵子结合,阻止转录;色氨酸不足,阻遏物三维空间结构发生变化,不能与操纵子结合,操纵元开始转录;色氨酸浓度升高:色氨酸与阻遏物结合,空间结构发生变化,可与操纵子结合,阻止转录; 12.说明下例每种酵母菌单倍体细胞的交配型表型。 (1)一个基因型为MATaMATα交配型基因的重复突变体。 (2)一个MATa细胞中HMLα盒的缺失突变体。 答:(1)在这个重复突变体中,MATa基因可以通过HMLα沉默子的同源重组,而转变成MATα,形成2份MATα交配型基因,表现为交配型α。同样,突变体中MATα可通过HMRa盒的同源重组作用,转变成MATa交配型基因,表现为交配型a。 (2)交配基因型MATa向MATα的转变,必须通过细胞中HMLα盒的同源重组才能产生,而HMLα盒的缺失则阻断了这一过程,因此该交配型表型只能为a型。 13.举例说明激素对基因表达的调控作用。 答:
双翅目昆虫幼虫唾腺细胞内有巨大的唾腺染色体,在幼虫发育的不同阶段,一至数个横纹带发生疏松(puff),即染色质线高度松散。疏松区出现大量的新合成的mRNA,疏松区出现的时间和部位随着发育阶段而顺序消长。以果蝇唾腺染色体为例:三龄前期,第三染色体不出现疏松区;三龄后期,74区EF段、75区B段、78区D段出现疏松区;前蛹期,以上三个疏松区消失,71区C-E段出现疏松区;成蛹期,71区C-E段出现疏松区消失,74区EF段、75区B段又出现疏松区;以上说明74区EF段、75区B段基因与幼虫的蜕皮和化蛹有关。 74区EF段、75区B段在幼虫蜕皮时发生疏松是和幼虫体内分泌蜕皮激素有关。蜕皮激素是一种类固醇(steroid)化合物,由幼虫前胸腺分泌,传送到虫体各部分,引发74区EF段、75区B段的基因转录,导致幼虫蜕皮。胸腺结扎试验,说明了蜕皮激素对唾腺染色体的疏松区开启的作用。在三龄早期用尼龙绳将唾腺部分紧紧扎起,结果被结扎的前半部分受到蜕皮激素的作用,提前化蛹,而后半部分仍为幼虫。唾腺细胞检查发现,前半部分唾腺细胞中第三染色体上74区EF段、75区B段、78区D段出现疏松,而后半部分唾腺细胞中第三染色体上相应部位没有出现疏松。说明蜕皮激素引起这些部位基因的活性。 类固醇是疏水性强的化合物,可经扩散通过质膜进入细胞。在细胞内类固醇与其受体结合成二聚体,这种二聚体一旦与目的基因启动子结合,就可直接启动目的基因的转录。
第九章基因工程和基因组学 本章习题 1.什么是遗传工程?它在理论上和实践上有什么意义? 答:遗传工程是将分子遗传学的理论与技术相结合,用来改造、创建动物和植物新品种、工业化生产生物产品、诊断和治疗人类遗传疾病的一个新领域。 广义的遗传工程包括细胞工程、染色体工程、基因工程、细胞器工程等。狭义的遗传工程即是通常讲的基因工程。本章只涉及狭义的遗传工程,即基因工程。 理论意义:遗传工程(基因工程)中的DNA重组主要是创造自然界中没有的DNA分子的新组合,这种重组不同于精典遗传学中经过遗传交换产生的重组。 实践意义:遗传工程(基因工程)技术的建立,使所有实验生物学领域产生巨大的变革。在工厂化生产药品、疫苗和食品;诊断和治疗遗传疾病;培养转基因动植物等方面都有非常重大的意义,即基因工程技术已广泛用于工业、农业、畜牧业、医学、法学等领域,为人类创造了巨大的财富。(详见第10题)。 2.简述基因工程的施工步骤。 答:基因工程的施工由以下这些步骤: ⑴.从细胞和组织中分离DNA; ⑵.媚苁侗鹛匾霥NA序列的限制性核酸内切酶酶切DNA分子,制备DNA片段; ⑶.将酶切的DNA片段与载体DNA(载体能在宿主细胞内自我复制连接),构建重组DNA分子; ⑷.将重组DNA分子导入宿主细胞,在细胞内复制,产生多个完全相同的拷贝,即克隆; ⑸.重组DNA随宿主细胞分裂而分配到子细胞,使子代群体细胞均具有重组DNA分子的拷贝; ⑹.从宿主细胞中回收、纯化和分析克隆的重组DNA分子; ⑺.使克隆的DNA进一步转录成mRNA、翻译成蛋白质,分离、鉴定基因产物。 3.说明在DNA克隆中,以下材料起什么作用。 (1)载体;(2)限制性核酸内切酶;(3)连接酶;(4)宿主细胞;(5)氯化钠 答:⑴.载体:经限制性酶酶切后形成的DNA片段或基因,不能直接进入宿主细胞进行克隆。一个DNA片段只有与适合的载体DNA连接构成重组DNA后,在载体DNA的运载下,才可以高效地进入宿主细胞,并在其中复制、扩增、克隆出多个拷贝。可作为DNA载体的有质粒、噬菌体、病毒、细菌和酵母人工染色体等。 ⑵.限制性核酸内切酶:限制性核酸内切酶是基因工程的基石。在细菌中这些酶的功能是降解外来DNA分子,以限制或阻止病毒侵染。这种酶能识别双链DNA分子中一段特异的核苷酸序列,在这一序列内将双链DNA分子切断。 ⑶.连接酶:将外源DNA与载体相连接的一类酶。 ⑷.宿主细胞:能使重组DNA进行复制的寄主细胞。 ⑸.氯化钠:主要用于DNA提取。在pH为8左右的DNA溶液中,DNA分子是带负电荷的,加入一定浓度的氯化钠,使钠离子中和DNA分子上的负电荷,减少DNA分子之间的同性电荷相斥力,易于互相聚合而形成DNA钠盐沉淀。另外,氯化钠也是细菌培养基的成分之一。
4.有一个带有氨苄青霉素和四环素抗性的质粒,在其四环素抗性基因内有一个该质粒惟一的EcoRI酶切点,今欲用EcoRI位点克隆果蝇DNA,构建一个基因库,连接的产物转化大肠杆菌菌株DH5,试问:⑴.在培养基中加入哪一种抗生素用于选择阳性克隆?⑵.对哪一种抗生素有抗性的质粒携带外源果蝇DNA片段?⑶.如果有的克隆可抗两种抗生素,如何解释? 答:⑴.在培养基中加入四环素结合影印法可用于选择阳性克隆。 ⑵.对氨苄青霉素有抗性的质粒携带外源果蝇DNA片段。 ⑶.这种克隆是没有受到EcoRI酶解的原始质粒或这些克隆都是自连形成的非重组体。 5.在构建一个真核生物核DNA库时,需要考虑哪些因素? 答:核基因库是将某一生物的全部基因组DNA酶切后与载体连接构建而成的。通常方法是,尽量提取大分子量的核DNA,用限制性酶酶切后,分离选择具有一定长度(大于15kb)的DNA片断,与适宜的载体连接构成重组DNA分子,根据所用的载体,选择相应的宿主细胞用于克隆。若载体是质粒,则将连接的重组DNA分子转化感受态细胞,收集所有的菌落即成为质粒基因库。如果载体是噬菌体或粘粒(cosmid),则将重组DNA分子体外包装成噬菌体后,感染细菌细胞,将所得到的所有重组噬菌体集中即是基因库。如果载体为BAC或YAC,将重组人工染色体导入相应的宿主细胞,收集得到的所有细胞即成为基因库。 真核生物的核DNA大,因此在构建核基因库时,通常要选择能够接受较大片段的载体,以减少克隆数量。若构建的基因库是以分离结构基因为主要目的的,通常选用λEMBL,λGEM,或粘粒。而那些将用于基因组作图和分析的基因库,则多选择BAC或YAC为载体。 6.根据下列凝胶电泳分析的结果,构建一个限制性酶图谱,并表明酶切位点及片段的碱基数,片段总长度为1300bp。电泳分析结果如下:内切酶DNA片段长度I350,950II200,1100I和II150,200,950 答:限制性酶切图谱从左到右是,200个碱基对位置是酶II的切点;350个碱基对位置是酶I的切点;图谱总长是1300个碱基对。 7.在下列6种限制性酶图谱中,有一种排列方式与凝胶电泳的带型是一致的。3种酶分别是:E:EcoRI、N:NcoI、A:AatII。
试回答: ⑴.根据电泳中DNA带型,选择正确的图谱并说明原因。 ⑵.在将这块凝胶转移后进行Southern杂交分析,带星点的是与pep基因杂交的信号带,说明pep在图谱中的位置。 答:⑴.从上到下的第五条应该为正确的图谱,因为经过上述三种酶切后,与左面的电泳图完全一致。 ⑵.根据Southern结果和酶切图的位置,pep应该在第五条图谱的3与4之间。 8.简述将除草剂基因转移到植物基因组的过程。 答:以农杆菌介导为例,说明这一过程。 ⑴.在无菌的组织培养下,从植物体的种子或无性器官建立高效的再生体系; ⑵.依据植物的种类,选择合适的质粒载体,将抗除草剂的基因连接到载体上,再将质粒引进根癌农杆菌; ⑶.植物的再生组织与上述农杆菌共同培养; ⑷.经过农杆菌感染的组织在含除草剂的培养基中进行选择; ⑸.抗除草剂的组织再生植株; ⑹.再生植株在温室进行抗除草剂试验; ⑺.有性繁殖的种类还要进行自交、回交测定和纯化。 9.简述基因组遗传图谱与物理图谱的异同。 答:遗传图谱的构建是根据任一遗传性状(如已知的可鉴别的表型性状、多型性基因位点、功能未知的DNA标记)的分离比例,将基因定位在基因组中。因此,遗传图谱是根据等位基因在减数分裂中的重组频率,来确定其在基因组中的顺序和相对距离的。物理图谱的构建不需要检测等位基因的差异,它既可以利用具有多型性的标记,也可以利用没有多型性的标记进行图谱构建,它将标记直接定位在基因库中的某一位点。 实际上这两种途径都需要利用分子遗传学的技术和方法。尽管这两种图谱是分别构建的,但是它们可以相互借鉴、互为补充,作为基因组图谱利用。 构建物理图谱的原因是:遗传图谱的分辨率有限、遗传图谱的精确性不高。
10.简述基因工程在工、农、医三方面的成就及发展前景。 答:基因工程在工业上的应用主要是生产医药产品,最典型的例子是通过细菌生产胰岛素,治疗糖尿病。到目前通过细菌已经生产了表皮生长因子、人生长激素因子、干扰素、乙型肝炎工程疫苗等10多种医药产品。 基因工程在农业上的应用:以转基因植物为标志的植物基因工程已经培养出许多抗除草剂、抗虫、抗病、抗逆的优良品种和品系,如在全世界范围内大量推广应用的抗除草剂的大豆、抗棉铃虫的棉花等。通过转基因羊大量表达人类的抗胰蛋白酶;克隆动物的成功,可以挽救濒危的稀有动物。 基因工程在医学上主要是用于遗传疾病的诊断、基因的治疗方面。 基因工程具有巨大和广泛的发展前景,将渗透到人类生活的各个方面。可以创造出营养价值更高、保健作用更好、抗逆性更强的植物种类;转基因动物的进展,可以生产出多种类的用于人类遗传性疾病治疗的药物;人类基因组计划的完成和基因定位的发展、尤其是核酸分子杂交原理和方法与半导体技术结合而发展起来的DNA芯片技术的出现和完善,将在人类遗传疾病的诊断和治疗等方面发挥重要作用。
第十章基因突变 本章习题 1.举例说明自发突变和诱发突变、正突变和反突变。 答:通过外界环境条件的自然作用,或由植物体内生理生化变化而发生的突变,称为自发突变。如寿星桃是由正常的桃突变而来(DwDw-Dwdw-dwdw)。自然条件下,水稻的矮生型、棉花的短果枝、玉米的糯性胚乳等性状,都是基因自发突变的结果。 人为地利用物理、化学或其它因素影响而发生的突变称为诱发突变。如辐射诱发引起的苹果短枝型变异、人工诱发樱桃紧凑型变异。 基因突变像许多生物化学反应过程一样是可逆的,即显性基因A可以突变为隐性基因a,而隐性基因a也可突变为显性基因A。前者通常称为正突变,后者称为反突变或回复突变。例如水稻有芒基因A可以突变为无芒基因a,而无芒基因a也可突变为有芒基因A。 2.什么叫复等位基因?人的ABO血型复等位基因的遗传知识有什么利用价值? 答:位于同一基因位点上的各个等位基因在遗传学上称为复等位基因。复等位基因并不存在于同一个体(同源多倍体是例外),而是存在于同一生物类型的不同个体里。 人的ABO血型就是蒊A、IB和i三个复等位基因决定着红细胞表面抗原的特异性。其中,IA基因、IB基因分别对i基因为显性,IA与IB为共显性。根据ABO血型的遗传规律可进行亲子鉴定等。 3.何为芽变?在生产实践上有什么价值? 答:芽变是体细胞突变的一种,突变发生在芽的分生组织细胞中。当芽萌发长成枝条,并在性状上表现出与原类型不同,即为芽变。 芽变是植物产生新变异的丰富源泉,它既可为杂交育种提供新的种质资源,又可从中选出优良新品种,是选育品种的一种简易而有效的方法。全世界有一半苹果产量来自于芽变,如品种:元帅、红星、新红星、首红、超首红。 4.为什么基因突变大多数是有害的? 答:大多数基因的突变,对生物的生长和发育往往是有害的。因为现存的生物都是经历长期自然选择进化而来的,它们的遗传物质及其控制下的代谢过程,都已经达到相对平衡和协调状态。如果某一基因发生突变,原有的协调关系不可避免地要遭到破坏或削弱,生物赖于正常生活的代谢关系就会被打乱,从而引起程度不同的有害后果。一般表现为生育反常,极端的会导致死亡。5.有性繁殖和无性繁殖、自花授粉和异花授粉与突变性状表现有什么关系? 答:有性繁殖植物:性细胞发生显性突变,则在后代中立即表现;如果是隐性突变,后代自交也可以得到纯合的突变体。体细胞发生显性突变,则以嵌合体形式存在;体细胞
发生隐性突变,不能立即表现,如要使它表现则需要把隐性突变体进行有性繁殖。 无性繁殖植物:体细胞显性突变后,形成嵌合体,用嵌合体进行无性繁殖,可以得到表现各种变异的嵌合体,也可能得到同质突变体;发生隐性突变则无法通过无性繁殖使之得到表现。 自花授粉植物:一般自花授粉植物突变频率低,遗传上较稳定,但是突变后容易表现,容易被检出。 异花授粉植物:异花授粉植物突变频率相对较高,但是突变后不容易被检出。因为显性突变成杂合状态存在,隐性突变大多被显性基因遮盖而不表现,只要在自交时基因型纯合,才能表现。 6.突变的平行性说明什么问题,有何实践意义? 答:亲缘关系相近的物种因遗传基础比较近似,往往发生相似的基因突变。这种现象称为突变的平行性。根据这个特点,当了解到一个物种或属内具有哪些变异类型,就能够预见到近缘的其它物种或属也可能存在相似的变异类型,这对于人工诱变有一定的参考意义。 7.利用花粉直感现象测定突变频率,在亲本状态配置上应该注意什么问题? 答:一般应该用隐性纯合体作母本,用显性纯合体经诱变处理的花粉作父本进行杂交。 8.在高秆小麦田里突然出现一株矮化植株,怎样验证它是由于基因突变,还是由于环境影响产生的? 答:如果是在苗期发现这种情况,有可能是环境条件如土壤肥力、光照等因素引起,在当代可加强矮化植株与正常植株的栽培管理,使其处于相同环境条件下,观察它们在生长上的差异。如果到完全成熟时,两者高度表现相似,说明它是不遗传的变异,由环境影响引起的;反之,如果变异体与原始亲本明显不同,仍然表现为矮秆,说明它可能是遗传的变异。然后进行子代比较加以验证,可将矮化植株所收种子与高秆小麦的种子播种在相同的环境条件下,比较它的后代与对照在株高上的差异。如矮化植株的种子所长成的植株仍然矮化,则证明在高秆小麦田里出现的一株矮化植株是由于基因突变引起的。 9.试述物理因素诱变的机理。 答:本章所指的物理因素只限于各种电离辐射和非电离辐射。 电离辐射包括α射线、β射线和中子等粒子辐射,还包括γ射线和X射线等电磁辐射。电离辐射能使构成基因的化学物质直接发生电离作用。轻者造成基因分子结构的改变,产生突变了的新基因,重者造成染色体的断裂,引起染色体结构的畸变。 本章所指的非电离辐射就是紫外线。紫外线造成基因分子链的离析。分子链已经离析的基因在重新组合的时候,有可能发生差错而出现基因突变。紫外线特别作用于嘧啶,使得同链上邻近的嘧啶核苷酸之间形成多价的联合。最通常的结果是促使胸腺嘧啶联合成二聚体;或是将胞嘧啶脱氨成尿嘧啶,或是将水加到嘧啶的C4、C5位置上成为光产物。它可以削弱C与G之间的氢键,使DNA链发生局部分离或变性。
10.试用红色面包霉的生化突变试验,说明性状与基因表现的关系。 答:射线照射后的分生孢子可诱发突变,让诱变过的分生孢子与野生型交配,产生分离的子囊孢子,放入完全培养基里培养生长(基本培养基上只有野生型能够生长,突变型均不能生长),鉴定是否突变: ⑴.取出完全培养基中各组分生孢子,分别于基本培养基上,如果能够生长,说明仍与野生型一样,没有突变;如不能够生长,说明发生了变异; ⑵.把确定为突变型的各组材料,分别培养于加入各种物质的基本培养基中,如某一培养基上能生长,就说明控制合成加入物质的这种基因发生了突变; ⑶.如在上步2中确定为缺乏维生素合成能力的突变型,再进一步在培养基中分别加入各种维生素分别培养这种突变型,如果其中一个能生长,则说明是控制该个维生素合成的基因发生了突变。 上述生化突变的研究,清楚地说明基因控制性状,并非基因直接作用于性状,而是通过一系列生化过程来实现的。
第十一章细胞质遗传 本章习题 1、什么叫细胞质遗传?它有哪些特点?试举例说明之。 答:细胞质遗传指由细胞质内的遗传物质即细胞质基因所决定的遗传现象和规律,又称非染色体遗传、非孟德尔遗传、染色体外遗传、核外遗传、母体遗传。 细胞质遗传的特点:⑴.遗传方式是非孟德尔式的;杂交后一般不表现一定比例的分离。⑵.正交和反交的遗传表现不同;F1通常只表现母体的性状,故又称母性遗传。⑶.通过连续回交能将母本的核基因几乎全部置换掉,但母本的细胞质基因及其所控制的性状仍不消失。⑷.由附加体或共生体决定的性状,其表现往往类似病毒的转导或感染。 举例:罗兹(RhoadesM.M.)报道玉米的第7染色体上有一个控制白色条纹的基因(ij),纯合的ijij植株叶片表现为白色和绿色相间的条纹。以这种条纹株与正常绿色进行正反杂交,并将F1自交其结果如下:当以绿色株为母本时,F1全部表现正常绿色与非绿色为一对基因的差别,纯合隐性(ijij)个体表现白化或条纹,但以条纹株为母本时,F1却出现正常绿色、条纹和白化三类植株,并且没有一定的比例,如果将F1的条纹株与正常绿色株回交,后代仍然出现比例不定的三类植株,继续用正常绿色株做父本与条纹株回交,直至ij基因被全部取代,仍然没有发现父本对这个性状的影响,可见是叶绿体变异之后的细胞质遗传方式。 2、何谓母性影响?试举例说明它与母性遗传的区别。 答:由于母本基因型的影响,使子代表现母本性状的现象叫做母性影响,又叫前定作用。 母性影响所表现的遗传现象与母性遗传十分相似,但并不是由于细胞质基因组所决定的,而是由于核基因的产物在卵细胞中积累所决定的,故不属于母性遗传的范畴。 举例:如椎实螺外壳的旋转方向有左旋和右旋,这对相对性状是母性影响。把这两种椎实螺进行正反交,F1外壳的旋转方向都与各自的母体相似,成为右旋或为左旋,但其F2却都有全部为右旋,到F3世代才出现右旋和左旋的分离。这是由一对基因差别决定的,右旋(S+)对左旋(S)为显性,某个体的表现型并不由本身的基因型直接决定,而是由母体卵细胞的状态所决定,母本卵细胞的状态又由母本的基因型所决定。F1的基因型(S+S)决定了F2均为右旋,而F2的三种基因型决定了F3的二种类型的分离,其中S+S+和S+S的后代为右旋,SS后代为左旋。 -******3、如果正反交试验获得的F1表现不同,这可能是由于⑴.性连锁;⑵.细胞质遗传;⑶.母性影响。你如何用试验方法确定它属于哪一种情况? 答:正反杂交获得的F1分别进行自交或近亲交配,分析F1和F2性状分离与性别的关系,如群体中性状分离符合分离规律,但雌雄群体间性状分离比例不同者为性连锁;若正交F1表现与母本相同,反交不同,正交F1与其它任何亲本回交仍表现为母本性状者,并通过连续回交将母本的核基因置换掉,但该性状仍保留在母本中,则为细胞质遗传。若F1表现与母本相似,而自交后F2表现相同,继续自交其F3表现分离,且符合分离规律,则为母性影响。 4、细胞质遗传的物质基础是什么?
答:所有细胞器和细胞质颗粒中的遗传物质均为细胞质遗传的物质基础。细胞器基因组包括:线粒体基因组、叶绿体基因组、动粒基因组、中心粒基因组、膜体系基因组等;非细胞器基因组包括细胞共生体基因组、细胞质粒基因组等。 5、细胞质基因与核基因所何异同?二者在遗传上的相互关系如何? 答:共同点:虽然细胞质基因在分子大小和组成上与核基因有某些区别,但作为一种遗传物质,在结构上和功能上与核基因有许多相同点:⑴.均按半保留复制;⑵.表达方式一样:DNA-mRAN核糖体-蛋白质;⑶.均能发生突变,且能稳定遗传,其诱变因素亦相同。 不同点:细胞质基因突变频率大,具有较强的定向突变性;正反交不一样,基因通过雌配子传递;基因定位困难;载体分离无规律,细胞间分布不均匀;某些基因有感染性。而核基因突变频率较小,难于定向突变性;正反交一样,基因通过雌雄配子传递;基因可以通过杂交方式进行定位;载体分离有规律、细胞间分布均匀;基因无感染性。 遗传学中通常把染色体基因组控制的遗传现象和遗传规律称为核遗传,把细胞质基因所决定的遗传现象和遗传规律称为细胞质遗传,两者在遗传上相互协调和制约,反映了核与质两个遗传体系相互依存和联系的统一关系。一般情况下,核基因在遗传上处于主导的地位,但在某些情况下表现出细胞质基因的自主遗传作用。 6、试比较线粒体DNA、叶绿体DNA和核DNA的异同? 答:与核DNA相比,线粒体DNA和叶绿体DNA具有某些特点,其中: 线粒体DNA的特点:⑴.线粒体DNA是裸露的双链分子,一般为闭合环状结构,但也有线性的;⑵.线粒体DNA分子量为60×106,长度为10~30mm;⑶.线粒体DNA与原核生物的DNA一样,没有重复序列;⑷.线粒体DNA浮力密度比较低;⑸.线粒体DNA碱基成分中G和C有含量比A和T少;⑹.线粒体DNA两条单链的密度不同,一条称重链(H链),另一条称轻链(L链);⑺.线粒体DNA单个拷贝非常小,在细胞总DNA中占的比例非常小。 叶绿体DNA的特点:⑴.叶绿体DNA也是双链分子,呈裸露的闭合环状结构;⑵.叶绿体DNA约为150kb;⑶.叶绿体DNA一般是多拷贝的;⑷.叶绿体DNA浮力密度因物种而异,但与核DNA有不同程度的差异;⑸.叶绿体DNA碱基成分因物种不同而不同,高等植物叶绿体DNA与核DNA相同,但藻类植物中的CG含量较核DNA低;⑹.与核DNA相比,叶绿体DNA缺少5-甲基胞嘧啶。 相同之处:三者都是遗传物质(DNA),能稳定遗传给子代,且以半保留方式复制,表达方式一样,也能发生突变,诱变因素相同。 7、植物雄性不育主要有几种类型?其遗传基础如何? 答:
植物雄性不育主要有核不育性、质核不育性、质不育性三种类型: ⑴.核不育型是一种由核内染色体上基因所决定的雄性不育类型,一般受简单的1-2对隐性基因所控制,纯合体表现雄性不育。也发现由显性雄性不育基因所控制的显性核不育,它只能恢复不育性,但不能保持不育性。 ⑵.质核不育型是由细胞质基因和核基因互作控制的不育类型,由不育的细胞质基因和相对应的核基因所决定的。当胞质不育基因S存在时,核内必须有相对应的一对(或一对以上)隐性基因rr存在时,个体才能表现不育,只有细胞质或细胞核存在可育基因时能够表现为可育。根据不育性的败育发生的过程可分为:孢子体不育,指花粉的育性受孢子体(植株)基因型所控制,与花粉本身所含基因无关;配子体不育,指花粉育性直接受雄配子体(花粉)本身的基因所决定。不同类型需特定的恢复基因。 ⑶.质不育型是由细胞质基因所控制的不育类型,只能保持不育性,但不能恢复育性。如IRRI运用远缘杂交培育的雄性不育系IR66707A(Oryzaperennis细胞质,1995)和IR69700A(Oryzaglumaepatula细胞质,1996)均具有异种细胞质源,其细胞质完全不同于目前所有的水稻雄性不育系。这两个不育系属于细胞质型不育系,故其不育性都只能被保持而不能被恢复。 8、一般认为细胞质的雄性不育基因存在于线粒体DNA上,为什么? 答:⑴.在20世纪60年代已发现玉米不育株的线粒体亚显微结构与保持系有明显的不同,从而推断雄性不育性可能与线粒体的变异有关; ⑵.分子生物学上发现,玉米的4种类型的细胞质,正常可育型N和不育型T、C、S。它们的线粒体DNA分子组成有明显的区别,而叶绿体DNA并没有明显的差别,且以这4种类型线粒体DNA作模板,在体外合成蛋白质,N型合成的蛋白质与其它3种均不相同,也推断存在于线粒体的基因组中; ⑶.已完成的玉米N型和T型的mtDNA限制性内切酶图谱表明,N型mtDNA分别含有6组和5组重复序列,但只有其中的2组是两种mtDNA所共有的。就限制性位点的分布及Southern杂交的结果看,N型和T型所特有的碱基序列分别为70kb(N)和40kb(T),其余500kb的序列相同,且已从T型mtDNA中分离出一个专化玉米T型胞质不育基因Furf13。 ⑷.Northernblotting分析表明,玉米正常株与C型不育株的mtDNA基因atpa,atpb和ckx*的转录产物的长度和数目不同,可能与C型雄性不育型的表现有直接关系。 ⑸.除玉米外,在甜菜,矮牵牛,水稻等植物中,也发现不育系与可育系在叶绿体DNA的结构上没有差异,但在线粒体上有明显差别。 9、如果你发现了一株雄性不育植株,你如何确定它究竟是单倍体、远缘杂交F1、生理不育、核不育还是细胞质不育? 答:如果这植株是单倍体,那么这植株矮小,并伴有其它不良性状,雌雄均为不育,PMC减数分裂中期大多数染色体为单价体;而如果这是远缘杂交F1植株就较高大,营养生长旺盛,PMC减数分裂中期染色体配对异常,雌雄配子均不育但雌性的育性强于雄性。生理不育是不可遗传的。核不育和细胞质不育均为雄性不育,雌配子正常可育,但核不育材料与其它材料杂交的F1一般为可育,F2的育性分离呈现出明显的规律性;而细胞质不育的杂交后代可以保持不育(父本为保持系)或恢复可育(父本为纯合恢复系)。因此,可以从植株性状的遗传、植株形态、花粉母细胞镜检和杂交试验进行确定和区分。 10、用某不育系与恢复系杂交,得到F1全部正常可育。将F1的花粉再给不育系亲本授粉,后代中出现90株可育株和270株不育株。试分析该不育系的类型及遗传基础。 答:该不育系类型为孢子体不育S(r1r1r2r2) S(r1r1r2r2)×N(R1R1R2R2)F1S(R1r1R2r2)全部正常可育 S(r1r1r2r2)×S(R1r1R2r2)F11可育(S(R1r1R2r2))+3不育(S(r1r1r2r2)+S(r1r1R2r2)+S(R1r1r2
r2)) 该不育系的不育类型的遗传基础为:其恢复基因有两个,存在基因互作。无论是杂交还是回交后代中,个体基因型中只有同时存在两个显性恢复基因时,才能起到恢复育性的作用。因此,在回交后代中出现1:3可育株与不育株的分离。 11、现有一个不育材料,找不到它的恢复系。一般的杂交后代都是不育的。但有的F1不育株也能产生极少量F2花粉,自交得到少数后代,呈3:1不育株与可育株分离,将F1不育株与可育亲本回交,后代呈1:1不育株与可育株的分离,试分析该不育材料的遗传基础。 答:该不育材料是由单显性基因控制的不育系,其基因型为(MSMS)。该材料与可育材料(msms)杂交,其杂合体后代均为不育。一旦F1个体中出现少量可育花粉,自交后代即产生3:1的不育株与可育株的育性分离。F1不育株与可育亲本回交,即产生1:1的育性分离。至于F1不育株出现少量可育花粉可能是该材料的育性表现受环境条件(日照和温度等)的影响,在某一特定条件下,杂合体表现为可育。
第十二章遗传与发育 本章习题 1、怎样理解细胞核和细胞质在个体发育中的相互作用? 答:在个体发育过程中,细胞核和细胞质是相互依存,不可分割的,但起主导作用的应该是细胞核。因为细胞核内的"遗传信息"决定着个体发育的方向和模式,为蛋白质(包括酶)的合成提供了模板(mRAN)以及其它各种重要的RNA,从而控制了细胞的代谢方式和分化程序。细胞质则是蛋白质合成的场所,并为DNA的复制、mRNA的转录以及tRNA、rRNA的合成提供原料和能量。例如,叶绿体的RuBp羧化酶的8个小亚基和8个大亚基虽然分别由核基因和叶绿体基因编码合成,但叶绿体的形成却是在核基因的控制之下。从另一方面看,细胞质中的一些物质又能调节和制约核基因的特性,使得相同的细胞核由于不同的细胞质的影响而导致细胞的分化。从细胞分化这一点说,细胞质的不等分裂起着重要的作用;没有细胞质的不等分裂,其后果只能是细胞数目的增加,不会有细胞的分化。 2、海胆的受精卵分裂成4个细胞时,将各个细胞分开,都能发育成小幼虫。而分裂成8个小细胞时,再把它们分开,为什么不能发育成小幼虫? 答:这是因为受精卵虽是单细胞,但它的细胞质内除显见的细胞器有分化外,还存在动物极和植物极、灰色新月体和黄色新月体等分化。这些分化的物质将来发育成什么组织和器官,大体上已经确定。海胆受精卵的第一、二次分裂,都是顺着对称轴的方向进行的。获得的4个卵裂细胞中的细胞质是完全的,而第三次卵裂方向与对称轴垂直,分裂的8个卵裂细胞的细胞质不均等,因此就不能发育成小幼虫。这说明了细胞质是胚胎发育所必需,而且一个细胞除核和各种细胞器外。其它的不同部分对个体发育也能产生不同的影响。 3、如何理解个体发育的阶段性和连续性? 答:在个体发育的过程中,各种性状的发育,从受精卵开始分裂时就开始了,随着个体发育时期的推进,相应的性状有序且有节奏的发生。这一过程实际上包括了一系列连续的发育阶段,这些阶段按预定的顺序依次接连发生。上一阶段的趋向完成,启动下一阶段的开始。在正常情况下,一个细胞(组织或器官)通常不再继续分化或转化为其它任何结构,个体发育的这种特性是由内外两种因素控制的。内在的因素也就是遗传的因素,是基因序列在不同时间上的选择性表达。外在因素则包括相邻细胞间或组织器官间以及外界环境条件的影响。 4、说明同形异位基因在个体发育中的作用。 答:同形异位基因控制着个体的发育模式,组织和器官的形成。同形异位基因编码一组转录因子,这类转录因子都有含有一段或几段十分保守的序列,形成一定空间结构,与特异的DNA序列结合,和RNA聚合酶及其它因子一起控制基因转录,即,同形异位基因是通过调控其它重要的形态及器官结构基因的表达(包括控制细胞分裂、纺锤体形成和取向、细胞分化等发育过程的基因)来控制生物发育及器官形成的。 5、何谓分化,去分化和再分化?植物细胞或组织怎样恢复它的全能性?
答:分化反映来自同一合子或遗传上同质的细胞转变为形态机能和化学构成上异质细胞的过程,是由一般变为特殊的现象,它可在细胞水平,组织水平和器官水平上表现出来。 去分化是指植物细胞或组织脱离其受发育阶段及所在组织器官环境的约束,成为游离状态,并且得到必需营养和激素,就可能经去分化而恢复其遗传全能性。 再分化指由脱分化状态的细胞再度分化形成另一种或几种类型的细胞过程。 植物细胞或组织经激素等因素或条件的诱导后可恢复其细胞的全能性,即脱分化。 6、简述利用植物细胞的全能性在作物育种上获得的成就? 答:利用植物细胞的全能性在作物育种上能获得以下几方面的成就: ⑴.胚珠培养,克服远缘杂交不亲和性,获得种间杂种。 ⑵.通过体细胞培养,创造体细胞无性系突变,从而创造出新的种质资源。 ⑶.通过原生体培养和体细胞杂交,创造出有性杂交难于获得的远缘杂交种。 ⑷.难过花粉或花培养,进行单倍体育种,提高育种效率,缩短常规育种年限。 ⑸.通过体细胞培养诱导体细胞胚,经包裹后生产人工种子。 ⑹.结合外源基因导入(农杆菌介导、基因枪等)技术,通过植物细胞全能性获得转基因植株。
第十三章数量性状的遗传 本章习题 1.解释下列名词:广义遗传率、狭义遗传率、近交系数、共祖系数、数量性状基因位点、主效基因、微效基因、修饰基因、表现型值、基因型与环境互作 广义遗传率:通常定义为总的遗传方差占表现型方差的比率。 狭义遗传率:通常定义为加性遗传方差占表现型方差的比率。 近交系数:是指个体的某个基因位点上两个等位基因来源于共同祖先某个基因的概率。 共祖系数:个体的近交系数等于双亲的共祖系数。 数量性状基因位点:即QTL,指控制数量性状表现的数量基因在连锁群中的位置。 主效基因:对某一性状的表现起主要作用、效应较大的基因。 微效基因:指一性状受制于多个基因,每个基因对表现型的影响较小、效应累加、无显隐性关系、对环境敏感,这些基因称为微效基因。 修饰基因:对性状的表现的效应微小,主要是起增强或减弱主基因对表现型的作用。 表现型值:是指基因型值与非遗传随机误差的总和即性状测定值。 基因型与环境互作:数量基因对环境比较敏感,其表达容易受到环境条件的影响。因此,基因型与环境互作是基因型在不同环境条件下表现出的不同反应和对遗传主效应的离差。 2.质量性状和数量性状的区别在哪里?这两类性状的分析方法有何异同? 答:质量性状和数量性状的区别主要有:①.质量性状的变异是呈间断性,杂交后代可明确分组;数量性状的变异则呈连续性,杂交后的分离世代不能明确分组。②.质量性状不易受环境条件的影响;数量性状一般容易受环境条件的影响而发生变异,而这种变异一般是不能遗传的。③.质量性状在不同环境条件下的表现较为稳定;而控制数量性状的基因则在特定时空条件下表达,不同环境条件下基因表达的程度可能不同,因此数量性状普遍存在着基因型与环境互作。 对于质量性状一般采用系谱和概率分析的方法,并进行卡方检验;而数量性状的研究则需要遗传学方法和生物统计方法的结合,一般要采用适当的遗传交配设计、合理的环境设计、适当的度量手段和有效的统计分析方法,估算出遗传群体的均值、方差、协方差和相关系数等遗传参数等加以研究。 3.叙述表现型方差、基因型方差、基因型×环境互作方差的关系。估计遗传协方差及其分量在遗传育种中有何意义? 答:表现型方差由基因型方差(VG)、基因型×环境互作方差(Ve)和环境机误方差()构成,即,其中基因型方差和基因型×环境互作方差是可以遗传的,而纯粹的环境方差是不能遗传的。 由于存在基因连锁或基因的一因多效,生物体的不同数量性状之间常存在不同程度的相互关连。在统计分析方法中常用协方差来度量这种相互关联的变异程度。由于遗传方差可以进一步区分为基因型方差和基因型×环境互作方差等不同的方差分量,故遗传协方差也可进一步区分为基因型协方差和基因型×环境互作协方差等分量。在作物遗传改良过程中,对某一性状进行选择时常会引起另一相关性状的变化,为了取得更好地选择效果,
并使一些重要的性状能够得到同步改良,有必要进行性状间的协方差即相关性研究。 如基因加性效应对选择是有效的,细胞质效应亦可通过母本得以传递,因此当育种的目标性状不易测定或遗传率较低、进行直接选择较难取得预期效果时,利用与其具有较高加性相关和细胞质相关的其它性状进行间接选择,则较易取得育种效果。显性相关则是控制性状的有关基因的显性效应相互作用而产生的相关性,杂交一代中表现尤为强烈,在杂种优势利用中可以加以利用。但这种显性相关会随着世代的递增和基因的纯合而消失,且会影响选择育种中早代间接选择的效果,故对于显性相关为主的成对性状应以高代选择为主。所以,进行各种遗传协方差分析更能明确性状间相关性的遗传本质,有利于排除环境因素对间接选择的影响,取得更好的选择效果,对于作物的选择育种具有重要的指导意义。 4.基于对数量性状遗传本质的理解,叙述数量性状的多基因假说的主要内容。 答:在遗传机制方面,数量性状受多基因控制,基因与基因间的关系错综复杂;数量基因的表达对环境条件的变化比较敏感,基因的作用与环境条件的影响混杂在一起。因此,数量性状的多基因假说的主要内容是: ⑴.数量性状受制于多对微效基因或称多基因的联合效应; ⑵.各对微效基因的效应相等而且是累加的,故又可称是累加基因; ⑶.各对基因对某一性状的效应微小,多基因不能予以个别的辨认,只能按性状的表现作为一个多基因体系进行研究; ⑷.微效基因之间无显隐性关系,一般用大写字母表示增效、小写字母表示减效作用; ⑸.微效基因对环境敏感,因而数量性状的表现易受环境的影响而发生变化; ⑹.微效基因具有多效性,除对数量性状起微效多基因的作用外,对其它性状有时也可能产生一定的修饰作用; ⑺.微效基因和主效基因均处于细胞核的染色体上,具有分离、重组、连锁等性质。 5.叙述主效基因、微效基因、修饰基因对数量性状遗传作用的异同。 答:主效基因、微效基因、修饰基因在数量性状遗传中均可起一定的作用,其基因表达均可控制数量性状的表现。但是它们对数量性状所起的作用又有所不同,主效基因的遗传效应较大,对某一数量性状的表现起着主要作用,一般由若干个基因共同控制该性状的遗传;修饰基因的遗传效应微小,主要是对主效基因起修饰作用,起增强或减弱主基因对表现型的作用;而微效基因是指控制数量性状表现的基因较多,而这些基因的遗传效应较小,它们的效应是累加的,无显隐性关系,对环境条件的变化较敏感,且具有一定的多效性,对其它性状有时也可能产生一定的修饰作用。 6.什么是普通遗传率和互作遗传率?他们在育种实践上有何指导意义? 答:遗传率是指基因型方差(VG)占表型总方差(Vp)的比值,它是衡量基因型变异和表型总变异相对程度的遗传统计量。遗传率反映了通过表型值预测基因型值的可靠程度,表明了亲代变异传递到子代的能力。同时也可以作为考查亲代与子代相似程度的指标。由于导致群体表现型产生变异的遗传原因可以进一步区分为由遗传主效应产生的普通遗传变异和由基因型×环境互作效应产生的互作遗传变异,故遗传率可以分解为普通遗传率和互作遗传率两个分量。其中普通遗传率是指由遗传主效应引起的那部分遗传率,一般指遗传方差占表现型方差的比率;互作遗传率是指由基因型×环境互作效应引起的那部分遗传率,一般指基因型×环境互作方差占表现型方差的比率。
育种实践表明,根据遗传率的大小可以决定不同性状的选择时期和选择方法,这对于改进育种方法,避免育种工作的盲目性和提高育种效果是很有效的。一些遗传率较高的性状,可在杂种的早期世代进行选择,收效比较显著:而对于遗传率较低的性状,则需要在杂种后期世代进行选择才能收到更好的效果。一般而言,当数量性状的基因型×环境互作效应越强,其互作遗传率就会越大,该性状的遗传表现就越易因环境而异,通过选择只能获得适应某一年份或某一特殊环境(如某一生态区域)的品种或组合;而基因型×环境互作效应小的性状则其普通遗传率就会越大,容易通过选择来改良育种材料的遗传组成,获得能够适应不同年份或不同环境的品种(组合)。故普通遗传率适用于不同环境条件下的选择,而互作遗传率则只适用于某一特定条件下的选择。某一年份或环境下的选择总效益,可以根据总的遗传率大小(普通遗传率+某一环境中的互作遗传率)进行预测和分析,以了解通过选择个体或个体群改良其基因型的准确性和选择效率。 7.什么是基因的加性效应、显性效应及上位性效应?它们对数量性状遗传改良有何作用? 答:基因的加性效应(A):是指基因位点内等位基因的累加效应,是上下代遗传可以固定的分量,又称为"育种值"。显性效应(D):是指基因位点内等位基因之间的互作效应,是可以遗传但不能固定的遗传因素,是产生杂种优势的主要部分。上位性效应(I):是指不同基因位点的非等位基因之间相互作用所产生的效应。 上述遗传效应在数量性状遗传改良中的作用:由于加性效应部分可以在上下代得以传递,选择过程中可以累加,且具有较快的纯合速度,具有较高加性效应的数量性状在低世代选择时较易取得育种效果。显性相关则与杂种优势的表现有着密切关系,杂交一代中表现尤为强烈,在杂交稻等作物的组合选配中可以加以利用。但这种显性效应会随着世代的递增和基因的纯合而消失,且会影响选择育种中早代选择的效果,故对于显性效应为主的数量性状应以高代选择为主。上位性效应是由非等位基因间互作产生的,也是控制数量性状表现的重要遗传分量。其中加性×加性上位性效应部分也可在上下代遗传,并经选择而被固定;而加性×显性上位性效应和显性×显性上位性效应则与杂种优势的表现有关,在低世代时会在一定程度上影响数量性状的选择效果。 8.什么是基因的加性×环境互作效应、显性×环境互作效应及上位性×环境互作效应?它们对数量性状遗传改良作用与基因的遗传主效应有何异同? 答:
加性×环境互作效应(AE):是指基因加性效应与环境互作产生的遗传效应,是一部分可以在上下代传递、并加以固定的遗传效应,但会因环境条件的变化而产生较大差异。 显性×环境互作效应(DE):是指基因显性效应与环境互作产生的遗传效应,是一部分可以遗传、但不能固定的遗传效应,主要与杂种的优势表现有关,这部分效应也会因环境的变化而异。 上位性×环境互作效应(IE):是指基因上位性效应与环境互作产生的遗传效应,在不同环境中会有较大差异。其中加性×加性上位性互作效应部分经选择可被固定;而加性×显性上位性互作效应和显性×显性上位性互作效应与杂种优势的表现有关,在低世代时会在一定程度上影响数量性状的选择效果。 不同的环境互作效应与遗传主效应的作用一样,对数量性状的遗传改良起着重要作用,可以影响数量性状的表现和选择效果。但两者也有较大的差异,特别是一些受基因加性、显性或上位性等遗传主效应控制的数量性状在遗传改良中较为稳定,不同环境条件对这些数量性状的选择效果影响较小,通过选择容易获得适合不同年份或不同地点的育种材料或组合。相反,某一数量性状的基因型与环境互作效应越强,该性状的遗传表现就越容易受到环境变化的影响,通过选择一般可以获得适合某一特定年份或某一特定环境的育种材料。因此不同环境下的基因型稳定性对作物种子品质育种目标的制定非常重要。 9.以下是陆地棉4个亲本及其F1在8月9日和9月3日的平均单株成铃数的分析资料(1981,1985) 表1方差和协方差计算结果:方差8月9日9月3日协方差CovVA2.250*8.182**CA4.277*VD3.809*2.735*CD2.373VAE2.207*1.086CAE1.086VDE0.605*1.644*CDE-0.156Ve3.216*6.634*Ce3.853+VP12.087**20.280**CP11.433* 表2基于群体均值的F1平均优势的预测结果(%)组合8月9日9月3日HMHME1HME2HMHME1HME2[1×2]18.9*-7.620.7+4.22.5-2.1[1×3]23.7**12.1+3.1-0.422.6**-26.0**[1×4]19.5+6.25.60.619.0**-21.5**[2×3]41.4**8.821.316.2+8.4**10.5[2×4]50.1*1.029.735.110.927.2[3×4]39.3*14.912.111.130.8**-18.3** ⑴.估算8月9日和9月3日的普通广义遗传率、互作广义遗传率、普通狭义遗传率、互作狭义遗传率,说明这两个时期单株成铃数的遗传规律及其对选择育种的指导意义;⑵.根据F1普通平均优势和互作平均优势的预测结果,评价不同杂交组合的杂种优势利用的潜力。 答:⑴.估算8月9日和9月3日的普通广义遗传率、互作广义遗传率、普通狭义遗传率、互作狭义遗传率,说明这两个时期单株成铃数的遗传规律及其对选择育种的指导意义; 8月9日单株成铃数的遗传率分量: 普通广义遗传率
互作广义遗传率 普通狭义遗传率 互作狭义遗传率 9月3日单株成铃数的遗传率分量: 普通广义遗传率 互作广义遗传率 普通狭义遗传率 互作狭义遗传率 从表中可以看出,除了9月3日单株成铃数的加性互作方差(VAE)外,其它表型方差(VP)、机误方差(Ve)以及遗传方差分量(包括加性方差(VA)、显性方差(VD)、加性互作方差(VAE)和显性互作方差(VDE))均已达到显著水平,说明基因的加性效应、显性效应、加性互作效应、显性互作效应均可显著影响两年不同时期的单株成铃数。在所分析的遗传主效应中,两个日期的平均单株成铃数都是以遗传主效应(VA+VD)为主,分别占VP的50.12%和53.83%,说明单株成铃数的选择效果受环境条件变化的影响相对较小。由于不同时期的单株成铃数的VAE和VDE多数已达显著水平,故该性状除了受到遗传主效应外,还不同程度受控于环境互作效应的影响,特别以8月9日的加性互作效应和9月3日的显性互作效应表现的尤为明显。这部分互作效应是单株成铃数在不同环境中遗传表现有所差异的主要原因。 通过表1中遗传效应的分析,还可以发现两年中单株成铃数均是以基因加性主效应和加性互作效应(VA+VAE)为主,且具有较高的普通狭义遗传率和互作狭义遗传率(分别为36.9%和45.7%),这表明对该性状进行低世代选择可望取得较好的效果。 由于不同时期单株成铃数的机误方差(Ve
)已达显著水平,所以该性状的表现还受到环境机误或抽样误差的影响。但由于其值均较小,故单株成铃数主要受制于加性效应、显性效应的各种遗传主效应以及相应的环境互作效应。 协方差分析的结果表明,8月9号和9月3号的表型或加性协方差已达显著水平,说明这两个时期的表型或加性效应间存在着显著正相关,通过8月9号的单株成铃数选择有利于增加9月3号的单株成铃数。由于未测到显著水平的互作效应协方差(加性互作协方差CAE和显性协方差CDE),故上述相关性受环境条件的影响不大。 ⑵.根据F1普通平均优势和互作平均优势的预测结果,评价不同杂交组合的杂种优势利用的潜力: 表2结果表明,8月9日棉花F1植株单株成铃数的普通平均优势(HM)在6个组合中均达到了1、5或10%的正向显著水平,表明普通平均优势可以显著增加该时期棉花的单株成铃数;由于8月9日时期的单株成铃数的互作优势仅有2个组合(1×2的HME2和1×3的HME1)达到了10%显著水平,因此该时期F1植株的单株成铃数平均优势受环境条件的影响较小。在9月3日时期,仅有组合2×3的平均杂种优势达到了10%显著水平(),说明该时期不同组合的单株成铃数普通平均优势不强;但9月3日时期单株成铃数的互作优势多数组合已达显著水平,说明该时期棉花单株成铃数杂种优势表现容易受到环境变化的影响,其中1981年主要为正向互作杂种优势,1985年则表现为以负向互作杂种优势为主。 就不同组合而言,组合2×3和2×4在8月9日具有较大的普通平均优势,而正向互作平均优势都未达到显著水平,故这两个组合的单株成铃数优势表现较好、且不同年份的单株成铃数也具有较好的稳定性。特别是组合2×3在9月3日时期的单株成铃数普通平均优势和1985年互作平均优势也已达到正向显著水平,表明该组合在不同发育时期(8月9日和9月3日)的单株成铃数具有较好的杂种优势。虽然组合1×3、1×4和3×4在8月9日的杂种优势表现较好,但这三个组合9月3日的显著互作优势在不同年份表现相反,表明这些组合在不同年份的稳定性较差。
第十四章群体遗传与进化 本章习题 1.解释下列名词:孟德尔群体、基因库、基因型频率、等位基因频率、遗传漂变、生殖隔离、地理隔离、进化树、进化速率、分子进化钟。 孟德尔群体:通过个体间的相互交配的结果,孟德尔遗传因子可以各种方式从一代传递给另一代的群体称为孟德尔群体。该群体不是一些个体的简单集合体,而是在各个体间有相互交配关系的集合体。 基因库:是指一个群体中全部个体所共有的全部基因称为基因库。 基因型频率:任何一个遗传群体都是由它所包含的各种基因型所组成的,在一个群体内某特定基因型所占的比例就是基因型频率。 等位基因频率:是指一群体内特定基因座中某一等位基因占该基因座等位基因总数的比率,或称基因频率。 遗传漂变:在一个小群体内,每代从基因库抽样形成下一代个体的配子时,会产生较大的抽样误差,由这种误差引起群体等位基因频率的偶然变化,叫做随机遗传漂变,或简称遗传漂变。 生殖隔离:是指防止不同物种的个体相互杂交的环境、行为、机械和生理的障碍。生殖隔离可以分为两大类:①.合子前生殖隔离,能阻止不同群体的成员间交配或产生合子;②.合子后生殖隔离,是降低杂种生活力或生殖力的一种生殖隔离。这两种生殖隔离最终达到阻止群体间基因交换的目的。 地理隔离:是由于某些地理的阻碍而发生的,例如海洋、大片陆地、高山和沙漠等,使许多生物不能自由迁移,相互之间不能自由交配,不同基因间不能彼此交流。 进化树:采用物种之间的最小突变距离构建而成的一种树状结构,可以表示不同物种的进化关系和程度,也称为种系发生树。一般是当不同物种蛋白质的氨基酸差异进一步以核苷酸的改变来度量时可用最小突变距离表示。 进化速率:进化速率是指在某一段绝对时间内的遗传改变量,一般可用不同物种的蛋白质、DNA和mtRNA等大分子的差异来估算进化速率。 分子进化钟:利用不同物种的蛋白质、DNA和mtRNA等大分子的差异估算出的分子进化速率,进而可以推断不同物种进化分歧的时间。 2.什么是生物的进化?它和遗传学有什么关系? 答:
生物进化是指在不断变化的自然条件下,生物体通过遗传、变异和自然选择并在隔离等因素的作用下,由简单到复杂、低级到高级的不断演变,可从旧物种中产生新的物种。其中微观进化是指发生在一个种内的进化,宏观进化是指在物种以上水平的进化。 遗传学所研究的是生物遗传和变异的规律和机理,进化论所研究的是生物物种的起源和演变过程。每个物种一般具有相当稳定的遗传特性,但新种的形成和发展则有赖于可遗传的变异。遗传学的研究不仅可以明确质量性状和数量性状的遗传规律,而且可从分子、细胞、个体和群体各个不同水平认识了遗传和变异的实质,为认识和控制生物的进化提供了理论和实践的依据,阐明生物进化的根本原因和历史进程,是研究进化论问题的必要基础。近代分子遗传学的发展更使进化论从分子水平上得到进一步的了解,根据遗传学研究的结果、采用实验方法人工创造和综合新的物种和新品种。如采用远缘杂交和细胞遗传分析等方法,已能清楚地说明小麦、棉花、烟草和芸苔属等物种的进化过程。因此,生物的进化是群体在遗传结构上的变化,人类对生物进化的认识,可以通过遗传学的研究不断向前发展。 3.拉马克和达尔文对于生物的进化有什么不同的看法?他们的进化观点还存在哪些不合理性? 答:拉马克写了一本《动物学哲学》,提出用进废退和获得性遗传原理来解释生物的进化。他认为动物和植物生存条件的改变是引起遗传特性发生变异的根本原因。而外界环境条件对生物的影响主要有两种形式,对于植物和低等动物的影响是直接的,如水生毛茛生长在水面上的叶片呈掌状,而生长在水面下的叶片呈丝状;而对于具有发达神经系统的高等动物则是间接的,当外界环境条件改变时,首先引起动物习性和行为的改变,然后促使某些器官使用的加强或减弱。这样,由于用进废退和获得性遗传,生物逐渐得到发展。但没有认识到选择在新物种或新类型形成过程中的作用,仅认为动物和植物的生存条件的改变是引起遗传特性发生变异的根本原因。 达尔文把选择的作用提到首要地位,认为新种的形成是一个极缓慢的过程,必须经过长时期的选择和积累作用,才能够形成新的物种。达尔文还提出性状分歧来解释物种的形成过程,认为物种和变种并没有本质的区别。只是程度不同而已。不同变种之间的差异常常比各物种之间的差异要小些,而变种乃是孕育中或正在形成的物种。达尔文学说的核心是选择,而作为选择的材料则是种内个体间的微小差异。在自然条件下,对于比较适应于条件的个体微小差异的选择和积累,这就是自然选择。在生物科学中,从这种观点认识生物发展和进化的,称作达尔文主义。但该进化观点认为生物体是由体质和种质两部分构成,环境条件的变化只能影响体质、不能影响种质,这种把生物体绝对化地划分为种质和体质的结果是否定了获得性状遗传,仅认为选择作用是形成新物种或新类型的主导因素,未考虑到生物的进化可以是突变、基因重组、选择和隔离等不同因素单独或共同起作用,更未考虑到由环境引起的变异是不能遗传的这一情况。 4.什么是自然选择?在生物进化中的作用怎样? 答:在自然界,一个种或变种内普遍存在着个体差异和繁殖过剩,加上自然产生的一些变异更会加剧个体间的差异,其结果必然产生生存竞争,一些强者或能够更好适应当时环境的个体在竞争中就能够获胜而得以生存。一般生物在发展过程中,如果新的变异类型比其它类型更适应环境条件,就能繁殖更多的后代,逐渐代替原有类型而成为新的种。如果新产生的类型和原有类型都能生存下来,不同类型分布在它们最适宜的地域,成为地理亚种。反之,当新的类型不及原有类型,就会被淘汰。 达尔文的生存竞争认为由种内竞争所产生的自然选择,是解释物种起源和生物进化的主要动力。由于新种的形成是一个极缓慢的过程,这些变异必须经过长时期的自然选择和积累,才可能形成新的物种。所以自然选择是生物界进化的主导因素,而遗传和变异则是它作用的基础。 5.什么是遗传的平衡定律?如何证明? 答:
在一个完全随机交配的群体内,如果没有其它因素(如突变、选择、迁移、遗传漂变等)干扰,则等位基因频率及三种基因型频率始终保持一定,各代不变。此事实为德国医生魏伯格和英国数学家哈迪在1908年分别发现,故称为“哈迪-魏伯格定律”。 证明:设一群体的等位基因A1和A2的频率分别为p1和p2(p1+p2=1),3种基因型的频率分别是:A1A1A1A2A2A2 如果进行随机交配,这个群体就能达到平衡。因为这3种基因型产生配子频率是: A1: A2: 如果个体间的交配是随机时,配子间的结合也是随机的,于是可得到以下结果:雄配子及其频率雄配子及其频率A1:p1A2:p2A1:p1A2:p2A1A1:p1Xp1=A1A2:p1Xp2=p1p2A1A2:p1Xp2=p1p2A2A2:p2Xp2= 下代3种基因型的频率分别是:A1A1A1A2A2A2 这个频率是和上代3种基因型的频率完全一样,所以就这对基因而言,该群体已经达到平衡。 6.有那些因素影响基因频率? 答:在自然界和栽培条件下,有着许多可以影响群体遗传平衡的因素,如突变、选择、迁移和遗传漂变等。这些因素都是促使生物发生进化的原因,其中突变和选择是主要的,遗传漂变和迁移也有一定的作用。 7.突变和隔离在生物进化中起什么作用? 答:
基因突变对于群体遗传组成的改变或生物进化有两个重要的作用。第一,它供给自然选择的原始材料,没有突变,选择即无从发生作用;第二,突变本身就是影响等位基因频率的一种力量。如一对等位基因,当基因A1变为A2时,群体中A1频率就会逐渐减少、A2频率则逐渐增加。假如长期连续发生A1→A2的突变,最后这一群体中A1将完全被A2代替。这就是由于突变而产生的突变压。 遗传学研究的结果已表明,物种之间具有较大的遗传差异,往往涉及一系列基因的不同以及染色体数目和结构上的差别。为了保证物种的进化和一定的稳定性,在不同物种或群体之间,往往会有一定的隔离方法。主要包括生殖隔离和地理隔离等,可以达到阻止群体间基因交换之目的,最终形成新的物种,促进生物进化。因此,隔离是巩固由自然选择所累积下来的变异的重要因素,也是保障物种形成的最后阶段,在物种形成上是一个不可缺少的条件。 8.什么叫物种?它是如何形成的?有哪几种不同的形成方式? 答:物种是具有一定形态和生理特征、彼此可以自由交配并产生正常后代以及具有一定自然分布区域的生物类群,是生物分类、防止和进化的基本单元。 物种的形成:达尔文在1859年发表的《物种起源》中,提出自然选择和人工选择的进化学说。认为物种是在遗传、变异和自然选择以及隔离等因素的作用下,由简单到复杂、低级到高级逐渐进化的,从旧物种中产生新的物种。其中微观进化是指发生在一个种内的进化,宏观进化是指在物种以上水平的进化。因此,生物进化是群体在遗传结构上的变化。 物种的形成主要包括两种方式:一种是渐变式的,往往先形成亚种,旧的物种需在一个很长时间内逐渐累积变异而演变成为新的物种,这是物种形成的主要形式;另一种是爆发式,这种方式是在短期内以飞跃形式从一种物种变成另一种物种,不一定需要悠久的演变历史,一般也不经过亚种阶段,主要通过染色体的变异或突变以及远缘杂交和染色体加倍,在自然界的作用下逐渐形成新种,这是高等植物,特别是种子植物的形成中一种比较普遍的形式。 9.多倍体在植物进化中起什么作用? 答:多倍体是物种演化的一种重要因素。由多倍体化产生的新物种一般不需要较长的演变历史,旧物种生物体通过染色体加倍,在自然界的作用下经较短时间即可形成新种,这在一些显花植物中显得尤为明显,而且栽培植物中多倍体的比例要比野生植物多。因此,多倍体是快速形成物种的一种形式,在一代或几代内就可产生出多倍体个体。小麦、棉花、烟草和芸苔属等物种的进化过程证明多倍体现象在进化的历史上起了重要的作用。 10.下表是CCR5等位基因在亚洲不同人群中三种基因型人数的观察值(Martinson,1997)。⑴.计算各人群中的基因型频率和等位基因频率及其标准差;⑵.计算基因型频率和等位基因的95%置信区间;⑶.比较亚洲不同人群的CCR5等位基因频率有否差异,亚洲人群与欧洲人群的等位基因频率有否差异。群 体观 察 值人数(n)+/++/-Δ32/Δ32巴基斯坦343220菲律宾262600蒙古595900泰国10110010 答:⑴.计算各人群中的基因型频率和等位基因频率及其标准差:以巴基斯坦人群CCR5基因为例,计算基因型频率和等位基因频率及其标准差: +/+基因型频率估计和标准差:
+/Δ32基因型频率估计和标准差: Δ32/Δ32基因型频率估计和标准差: CCR5+的等位基因频率估计值和标准差: CCR5-Δ32的等位基因频率估计值和标准差: ⑵.计算基因型频率和等位基因的95%置信区间:以巴基斯坦人群为例,CCR5基因的基因型频率和等位基因频率的95%置信区间的估算如下: +/+基因型: +/Δ32基因型: Δ32/Δ32基因型: CCR5+的等位基因: CCR5-Δ32的等位基因: 同理可以分析得到其它人群的各种基因型频率和基因频率,其结果列表如下: 表亚洲不同人群中CCR5等位基因的频率估计值(标准差)群体观察值基因型频率估计值(标准差)
基因频率估计值(标准差)人数+/++/-Δ32/Δ32+/++/-Δ32/Δ32CCR5+CCR5-Δ32巴基斯坦3432200.9412(0.0403)0.0588(0.0403)0.0000(0.0000)0.9706(0.0202)0.0294(0.0202)菲律宾2626001.0000(0.0000)0.0000(0.0000)0.0000(0.0000)1.0000(0.0000)0.0000(0.0000)蒙古5959001.0000(0.0000)0.0000(0.0000)0.0000(0.0000)1.0000(0.0000)0.0000(0.0000)泰国101100100.9901(0.0099)0.0099(0.0099)0.0000(0.0000)0.9951(0.0049)0.0049(0.0049) 表亚洲不同人群中CCR5基因的基因型频率和等位基因频率的95%置信区间群体基因型频率等位基因频率+/++/-Δ32/Δ32CCR5+CCR5-Δ32上限下限上限下限上限下限上限下限上限下限巴基斯坦1.02020.86220.06190.05570.00000.00001.01020.93100.0690-0.0102菲律宾1.00001.00000.00000.00000.00000.00001.00001.00000.00000.0000蒙古1.00001.00000.00000.00000.00000.00001.00001.00000.00000.0000泰国1.00940.97080.0292-0.00940.00000.00001.00470.98550.0145-0.0047 ⑶.比较亚洲不同人群间、亚洲人群与欧洲人群间的CCR5等位基因频率有否差异。根据上述置信区间的结果,可以比较不同群体的等位基因频率差异性。由于菲律宾、蒙古和泰国人群的CCR5+基因频率的上下限均位于巴基斯坦人群的CCR5+基因频率置信区间(1.0102-0.9310)之间;而菲律宾与蒙古人群的CCR5+基因频率的上下限一样,且均在泰国人群的CCR5+基因型频率的置信区间(1.0047-0.9855)之间,故可以推断CCR5+基因频率在这4个国家间无显著差异。同理可以发现菲律宾和蒙古人群中CCR5-Δ32基因频率的上下限均为0,而泰国人群中CCR5-Δ32基因频率的置信区间为0.0145-0.0047,均介于巴基斯坦人群CCR5-Δ32基因频率的置信区间(0.0690-
-0.0102)内,因此这几个国家在CCR5-Δ32基因频率上的差异不显著。 亚洲和欧洲8个国家人群的置信区间表明,巴基斯坦人群的CCR5+基因频率的下限(0.9310)大于冰岛(0.9030)、英国(0.9144)的上限,所以CCR5+基因频率在亚洲巴基斯坦和欧洲冰岛、英国人群间存在着显著差异;希腊人群的CCR5+基因频率的置信区间(1.0025-0.9499)位于巴基斯坦的上下限之间、意大利的上限为0.9772处于巴基斯坦的置信区间(1.0102-0.9310)之间,两者之间均不存在显著差异。菲律宾和蒙古人群的CCR5+基因频率的上下限均为1.0000,显著大于冰岛(0.9030)、英国(0.9144)、意大利(0.9722)的上限,但与希腊人群(1.0025-0.9499)无显著差异。泰国人群的CCR5+基因频率的下限(0.9855)大于冰岛(0.9030)、英国(0.9144)或意大利(0.9772)的上限,故存在显著差异;但泰国(1.0047-0.9855)与希腊(1.0025-0.9499)人群间差异不显著。在CCR5-Δ32基因频率差异上,巴基斯坦人群的上限(0.0690)小于冰岛(0.0970)和英国(0.0856)下限,故存在显著差异;希腊的置信区间(0.0501-0.0000)位于巴基斯坦的置信区间(0.0690--0.0102)之间,两个国家无显著差异;意大利的置信区间(0.0871-0.0228)与巴基斯坦的置信区间(0.0690--0.0102)存在交叉,两者不存在显著差异。菲律宾和蒙古人群的CCR5-Δ32基因频率的上下限均为0.0000,小于冰岛(0.0970)、英国(0.0856)、意大利(0.0228)的下限,故与这些国家之间存在显著差异;但与希腊(0.0501-0.0000)差异不明显。泰国人群CCR5-Δ32基因频率的上限(0.0145)小于冰岛(0.0970)、英国(0.0856)和意大利(0.0228)的下限,因此与这些欧洲人群有着显著差异;但是泰国的上限(0.0145)位于希腊的置信区间(0.0501-0.0000)内,所以这两个国家的人群间无显著差异。'
您可能关注的文档
- 逻辑学(姜全吉)一到十一章课后习题答案.doc
- 逻辑学(第3版),课后练习题答案(部分),人大出版社.doc
- 逻辑学各章练习题(附答案).doc
- 逻辑学基础教程课后练习题部分参考答案2.doc
- 逻辑学导论章节练习题参考答案.doc
- 逻辑学导论练习题参考答案.doc
- 逻辑学导论陈波著第二版章节练习题参考答案及逻辑学导论综合练习100题参考答案.doc
- 逻辑学课后练习题答案.doc
- 遗传与进化课后习题答案.doc
- 遗传学答案.doc
- 遗传学课后习题答案.doc
- 遥感数字图像处理教程课后习题主要考点答案.doc
- 邓毛三课后习题答案.doc
- 邢台学院《电路》邱关源第五版课后习题答案详细解析.doc
- 邱关源,电路,第五版,物理学 答案 第六章.pdf
- 邱关源《电路》第5版课后习题答案1-8章.doc
- 邹理和《数字信号处理上》课后习题答案.pdf
- 郑杭生《社会学概论新修》复习题.doc
相关文档
- 施工规范CECS140-2002给水排水工程埋地管芯缠丝预应力混凝土管和预应力钢筒混凝土管管道结构设计规程
- 施工规范CECS141-2002给水排水工程埋地钢管管道结构设计规程
- 施工规范CECS142-2002给水排水工程埋地铸铁管管道结构设计规程
- 施工规范CECS143-2002给水排水工程埋地预制混凝土圆形管管道结构设计规程
- 施工规范CECS145-2002给水排水工程埋地矩形管管道结构设计规程
- 施工规范CECS190-2005给水排水工程埋地玻璃纤维增强塑料夹砂管管道结构设计规程
- cecs 140:2002 给水排水工程埋地管芯缠丝预应力混凝土管和预应力钢筒混凝土管管道结构设计规程(含条文说明)
- cecs 141:2002 给水排水工程埋地钢管管道结构设计规程 条文说明
- cecs 140:2002 给水排水工程埋地管芯缠丝预应力混凝土管和预应力钢筒混凝土管管道结构设计规程 条文说明
- cecs 142:2002 给水排水工程埋地铸铁管管道结构设计规程 条文说明